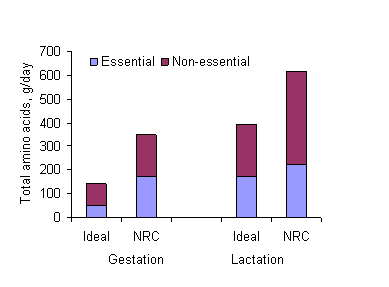UTA-TOSOLY - Finca Ecológica
Morario, Guapota, AA # 48, Socorro, Santander,
Santander del Sur, Colombia trpreston@mekarn.org
The original version of this paper was accepted for
publication in
CABI Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and
Natural Resources (http://www.cabi.org)
as of 12 September 2006
Abstract
The decreasing availability and increasing prices of
petroleum-based fuels is already having an impact on availability
of feed resources. Research to develop alternative sources of live
stock feed thus has a high priority. It is also important that the
alternative feed resources can be grown on the farm as this will:
(i) directly benefit the poorer farmers, who do not have cash
resources to purchase supplements from outside the farm; and (ii)
be an active response to the need to localize the farming system as
a defense against the decreasing availability and increasing prices
of petroleum-based fuels which will drive up the costs of
transport.
This review has shown that the leaves from shrubs such as
cassava and mulberry, from vegetables such as sweet potato and
cocoyam, together with water plants such as duckweed (Lemna
spp) and water spinach (Ipomoea aquatica), can be used
successfully in diets for pigs to replace at least half the protein
usually supplied as soya bean and fish meals.
It is strongly recommended that future research in this area
should be with basal diets that are low in protein and in cell
wall constituents, as this will facilitate: (i) increasing the
percentage of forage protein in the diet; and (ii) decreasing the
overall protein level in the diet in accordance with the concept of
the "ideal" protein.
Feed resources in the age of declining oil supplies
Conventional feed resources for pigs are cereal grains balanced
either by soybean meal or fish meal or combinations of the two.
These feed resources promise to follow the trends in fossil fuel,
namely a steady increase in the gap between demand and production
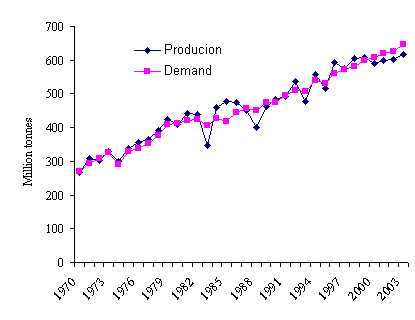
(Figure 1) and consequent increases in price.
Figure 1: World production and demand for
maize (Rameker 2004)
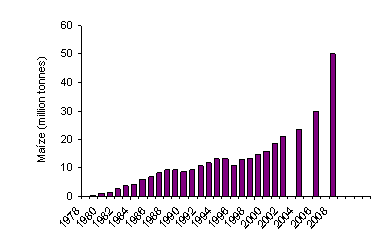
This is because cereal grain is increasingly being used as a
source of fuel via its conversion to ethanol (Figure 2).
Figure 2: Past and future trends for conversion of maize to
alcohol in USA
(Pearce Lyons and Bannerman 2001)
The sustainability of ethanol derived from biomass is presently
the subject of a fierce debate between: independent scientists
(Pimmental and Pazdek 2005) who calculate that it requires more
energy to produce than is contained in the resulting ethanol; and
government and industry representatives who argue that there is a
net increase of 30% relative to the energy costs of production.
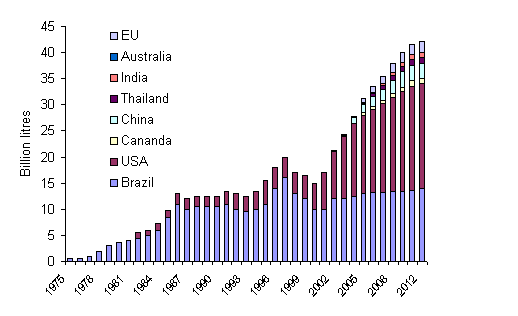
Nevertheless the reality is that in all parts of the developed
world and in advanced developing countries such as Brazil, China,
India and Thailand, new ethanol distilleries are being constructed
(Figure 3).
Figure 3: Global tendencies in production of
alcohol (Berg 2001)
Irrespective of the arguments over its sustainability, there is
not enough feedstock (principally cereal grains, sugar cane and
cassava roots) for ethanol production to completely replace oil as
a transport fuel on a world basis. Thus other solutions must be
found. One alternative is the decentralized production of
electricity by gasification of fibrous biomass and the use of the
electricity to charge the batteries in cars driven partially or
wholly by electric motors. This approach has the advantage of
generating much employment in rural areas through growing and
processing of the biomass.
Implications of ethanol and biomass gasification for live stock
production and especially pig production.
One obvious result of the increasing use of grain for alcohol
production is that its availability as live stock feed will
decrease while prices will increase. Thus there is a need to search
for alternatives. By contrast, the promotion of gasification will
create opportunities for such alternatives, in the form of crops
that produce both fibre and feed.
A secondary result of increasing fuel prices is to favor
de-centralized systems of production which are less dependent on
transport for the acquisition of inputs. This will also impact
strongly on past trends for pig production which have resulted in
large scale units (units with 1000 sows and followers are not
uncommon) that are completely dependent on importing / transporting
their feed supplies. Finally there are environmental issues such as
the pollution involved in the disposal of excreta in such large
operations and the difficulty (impossibility) of using efficiently
this resource as a fertilizer to replace fossil fuel-dependent
chemical fertilizers the prices of which will follow those of oil
and gas.
There is therefore high priority to develop alternative feeding
and management systems for pigs, as well as other live stock, which
can be sustained within their own land base. Apart from the
political and environmental benefits of reducing the dependence on
oil, and oil-related resources such as cereal grains, there will be
social benefits in that land-based farming systems will favor
smaller scale units in which all the family can participate, with
resultant increases in employment opportunities in rural areas.
Land-based farming systems
The questions to be addressed are:
What alternatives exist for development of pig feeding systems,
that use resources grown on the farm?
How should these resources be used in order to maximise the
benefits in the overall farming system?
An early hypothesis, which was proposed as the basis of a
research strategy for tropical latitudes, was that sugar cane could
be the basis of an integrated farming system in which the juice
expressed from the stalk in a simple 3-roll press could be used as
the energy source for pigs while the residual bagasse could be the
feedstock for gasification to produce electricity (Preston 1989).
The advantage seen in such a system was that the juice, composed
solely of 100% digestible sugars, would create opportunities for
using forages as the protein source, since the absence of fibre in
the juice would facilitate the intake of these fibrous feeds.
Unfortunately, the system depended on making efficient use of
the bagasse as a source of energy for gasification. This was shown
to be technically feasible (Preston T R and Lindgren A 1980,
unpublished data) but in an era of cheap oil was not economically
viable. So the hypothesis remained unproven and the proposed
research on using forages as the protein source was not
initiated
However, as indicated in the introductory paragraphs, the world
in 2006 is very different from what it was in 1989. Not only is the
need to replace oil energy with that from biomass a political issue
(Bush W, State of the Union speech 2 006), but the predicted
scarcity and high prices of cereal grains will make more attractive
the use of alternative sources of energy for live stock. The 1989
hypothesis can now be tested under favourable economic and
political conditions.
A basic feature of the original hypothesis was that in order to
use efficiently the protein in forages the basic energy source
should be low in fibre. There are a number of tropical feeds which
satisfy this condition:
The juice extracted from sugar cane stalks
The sap from the flower of the sugar palm tree (Borassus
flabelius)
The roots from cassava, taro, giant taro, cocoyam and sweet
potato
The fruit of banana and plantains
Broken rice
When the basal diet is derived from any of the above feeds, it
is possible to reduce the protein level, as when the protein is
derived mainly from the supplement, the amino acid balance is
better and approximates to the "ideal" protein (Wang and Fuller
1989). In this case, it has been shown that total protein levels
can be reduced by 30 to 40% compared with the case of diets based
on cereal grains (Figure 4).
Figure 4: Requirements for amino acids in sows fed a diet
with an "ideal" protein (Wang and Fuller 1989)
compared with the
traditional maize-soybean diet (NRC 1988)
Experiments with high levels of protein-rich foliages
Duckweed (Lemna spp.) (Figure 5)
One of the first experiments to evaluate diets in which all of
the protein was derived from forage was reported by Rodríguez
and Preston (1996). They used young pigs of a local Vietnamese
breed (Mong Cai) and crosses of Mong Cai with Large White, giving
them free access to sugar cane juice and fresh duckweed (protein
content of 35% in dry matter), which had been fertilized with
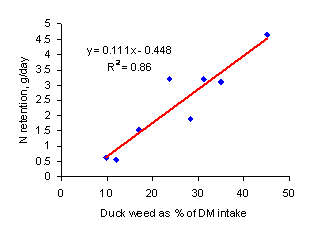
biodigester effluent. Nitrogen retention increased linearly
according to the proportion of duckweed in the diet (Figure 6).
Figure 5: Duckweed (Lemna
spp)
Figure 6: Relationship between N retention and
proportion of duckweed consumed by Mong Cai and Mong Cai crossbred
piglets offered a basal diet of sugar cane juice (Rodríguez
and Preston 1996)
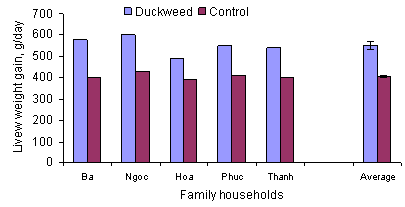
On-farm results confirmed the potential benefits from
the use of duckweed in pig diets (Figure 7).
Figure 7: Supplements of fresh duckweed increased growth
rates of pigs fed rice
byproducts on farms in Central Vietnam (Du Thanh Hang 1998)
Cassava (Manihot esculenta) leaves (Figure 8)
Many studies have evaluated use of cassava leaves as partial
replacement for soya bean meal and fish meals in conventional diets
for pigs (see review by Bui Ngu Phuc 2001). However, the levels
used rarely exceeded 15% in the diet, and the emphasis was mainly
on methods of reducing the risk of cyanide toxicity from cyanogenic
glucosides in the leaves. Sun-drying appeared to be more effective
than ensiling in this respect (Bui Huy Nhu Phuc et al 1996).
Figure 8: Cassava (Manihot esculenta)
Despite this concern for cyanide toxicity it is relevant to note
that there appear to have been no reported deaths of pigs from this
cause. A possible reason that has been suggested for this lack of
effect (R A Leng, personal communication) is that the enzymes
responsible for the release of HCN from the cyanogenic glucosides
are inactivated at the low pH in the pig stomach.
The uncertainly relating to possible risks of HCN toxicity led
Du Thanh Hang and Preston (2005) to compare processing of the fresh
cassava leaves by washing, chopping and washing; and chopping,
washing and wilting, and feeding them to pigs of 25 kg live weight
as supplements to a basal diet of cassava roots and rice bran. The
cassava leaves were readily consumed providing 38% of the dietary
DM and over 70% of the dietary protein with no effect of processing
method on total DM intake, which ranged from 27 to 31 g/kg live
weight. Levels of HCN were reduced slightly (16%) by washing and
substantially (82%) by wilting, resulting in intakes of HCN between
6.0 and 15 mg/kg live weight, levels considerably higher than the
range of 1.4 to 4.4 mg/kg live weight, previously reported as safe
to avoid toxicity (see Du Thanh Hang and Preston 2005). Following
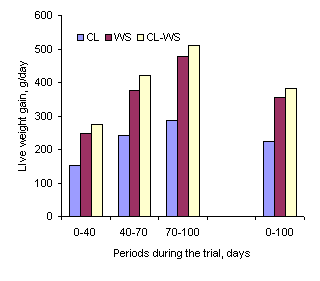
on from these findings, Chhay Ty and Preston (2005a,b) fed fresh
cassava leaves as the only supplement to a basal diet of broken
rice, comparing this treatment with fresh water spinach foliage or
a 50:50 mixture (DM basis) of the two vegetative sources. Feed
intakes, growth rates and DM digestibility coefficients were higher
when the water spinach was the only supplement or when it was mixed
with the cassava leaves, as compared with the cassava leaves alone
(Figure 9).
Figure 9: Growth rates of pigs (from 11 to 50 kg) fed
broken rice supplemented with fresh cassava leaves (CL),
water
spinach (WS) or a mixture of the two (CL-WS) (Chhay Ty and Preston
2005b)
Water spinach (Ipomoea aquatica) (Figure 10)
Water spinach grows equally well in the water or in soil. It is
traditionally consumed by people in SE Asia and appears to be
devoid of non-nutritional elements. Harvesting this plant from
lagoons fertilized with waste water from urban centres is an
important source of income for poor peoples in Vietnam, Cambodia
and Laos. An important feature of water spinach is its capacity to
yield high levels of biomass when fertilized with effluent from
biodigesters charged with pig manure (Kean Sophea and Preston
2001).
Figure 10: Water spinach (Ipomoea
aquatica)
Like the mulberry plant, consumed by silkworms, it was to be
expected that water spinach, consumed as a vegetable by humans,
would also have high nutritive value. Research in Cambodia confirms
that this is so. Basal diets of broken rice (8% protein in DM) in
which water spinach provided 50% of the dry matter and 70% of the
protein were highly digestible by pigs and supported higher N
retention than similar diets in which cassava leaves were the
supplementary protein source (Chhay Ty and Preston 2005a). Growth
rates of young pigs on these diets paralleled the N retention data
(Chhay Ty and Preston 2005b).
Mulberry (Morus alba) leaves (Figure 11)
The leaves of the Mulberry bush have long been used as substrate
for growth of the larvae of the silkworm. It could therefore be
expected that they would have potential as a protein source for
pigs.
Figure 11: Mulberry (Morus alba)
Chiev Phiny et al (2003) compared sun-dried leaves as
partial or complete replacements for rice bran and fish meal in a
basal diet of broken rice. Nitrogen retention appeared to be
improved with increasing levels of mulberry leaf meal in the diet,
when expressed as a percentage of N ingested or N digested. There
were no differences in digestibility or nitrogen retention when
fresh or sun-dried leaves were included at levels of 30% (DM basis)
in a diet in which the energy component was wheat bran.
Sweet potato (Ipomoea batatas) (Figure 12)
A very comprehensive study on sweet potato vines was done by Le
Van An (2004). He concluded that "the best options in terms of leaf
and stem production were a cutting interval of 20 days and a
defoliation of 50% of the total branches. Defoliation reduced tuber
production.
Figure 12: Sweet potato (Ipomoea batata)
There appear to be considerable differences, depending on
variety, in the content of crude protein and crude fibre in the dry
matter of the foliage of sweet potato, the former ranging from 26.5
to 32.5 % in leaves and from 10.4 to 14.1 % in stems (Woolfe 1992;
Ishida et al 2000; Le Van An 2004). For crude fibre the mean values
were 11.1 and 20.7 %, respectively (Woolfe 1992). It is therefore
important to separate leaves from stems when the aim is to maximize
the rate of inclusion of sweet potato foliage in pig diets.
According to Le Van An (2004) there are no major differences in
nutritive value between fresh, sun-dried or ensiled leaves. Adding
synthetic lysine (L) to a basal diet of rice by-products
supplemented with sweet potato leaves (SP) increased pig growth
rates to a level which did not differ from that on the positive
control diet containing fish meal (FM) and which were better than
on the diet supplemented with groundnut cake (GC). Daily live
weight gains of pigs on the FM, GC, SP and SPL diets were 542, 464,
482 and 536 g, respectively (P<0.05).
Giant Taro (Alocasia macrorrhiza) leaves
This plant appears to have been neglected by live stock
researchers despite the fact that it is widely distributed in
tropical latitudes, and possesses desirable characteristics such as
high protein (24% in DM) and low content of crude fibre (15% in DM)
(Gohl 1973). This author described the Giant Taro as a "herbaceous
plant with large heart-shaped leavc to 1.5 m long that contain
a milky juice. The wild variety has a pungent taste caused by
oxalate crystals which also cause the mouth to itch. Cultivated
forms do not have this property, and the leaves, stalks and black
rhizome can be fed to animals".
Interviews with farmers in Cambodia and Vietnam (Preston T R ,
unpublished data) failed to elucidate the meaning of "cultivated"
as opposed to "wild" forms. Most farmers indicated that they
traditionally "boiled" the leaves before feeding them to pigs as in
the fresh state the leaves were not readily consumed.
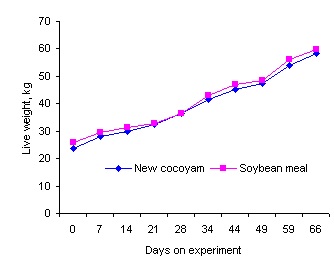
New Cocoyam (Xanthosoma sagittifolium) (Figure 13)
Figure 13: New cocoyam (Xanthosoma sagittifolium)
New Cocoyam, which appears to be native to South and Central
America, is similar in appearance to the Giant Taro (Alocasia
macrorrhiza). However, in contrast to the Giant Taro, the fresh
leaves of the "New Cocoyam" were found to be highly palatable to
pigs (Rodríguez et al 2006). At a level of 50% of substitution
of the soybean meal protein in a sugar cane juice diet for growing pigs,
the growth rates were the same as when the soybean meal provided
all the protein (Figure 14).
Figure 14: Growth curves of pigs fed a basal
diet of sugar cane juice
supplemented with soybean meal or a 50: 50 mixture (equal
protein basis)
of leaves of "New Cocoyam" and soybean meal
Conclusions
The greater part of the soybean meal and maize fed to pigs
throughout the world originates in the USA and Brazil. As transport
costs rise, driven by decreasing availability of cheap oil, this
inevitably will result in increased costs to farmers that have
become "addicted" to these feed resources as the basis of their pig
production systems.
Research to develop alternative sources of protein therefore has
a high priority. It is also important that the alternative feed
resources can be grown on the farm as this will: (i) directly
benefit the poorer farmers, who do not have cash resources to
purchase supplements from outside the farm; and (ii) be an active
response to the need to localize the farming system as a defense
against the decreasing availability and increasing prices of
petroleum-based fuels which will drive up the costs of transport.
This review has shown that the leaves from shrubs such as
cassava and mulberry, from vegetables such as sweet potato and
cocoyam, together with water plants such as duckweed (Lemna
spp) and water spinach (Ipomoea aquatica), can be used
successfully in diets for pigs to replace at least half the protein
usually supplied as soybean and fish meals.
The use of these vegetative protein sources is facilitated when
they are accompanied by energy sources that are low in both fibre
and protein. A low content of fibre in the energy source creates
"space" in the digestive tract of the pig for fibre present in the
leaves. In contrast to leaf proteins, which are mainly present as
enzymes having a balanced array of essential amino acids (Table 1),
the readily digestible energy in plants acts as a reserve supply
of nutrients to promote germination or recovery in growth after a
period of stress, such as happens in the dry season. With the
exception of rice, the protein associated with the energy in cereal
grains is also a store of nutrients but one in which the amino acid
array is usually imbalanced (eg: the deficiency of lysine in
maize). By contrast, the nutrient reserves in most tropical crops
(rice being the exception), contain little or no protein (eg: the
roots of cassava, tubers of sweet potato and taro, the fruit of
bananas and plantains, and the stalks of sugar cane). They also are
low in fibre. Thus there is potential for a natural synergism
between sources of energy and protein in tropical latitudes.
When energy sources low in protein (eg: sugar cane juice,
cassava roots, sweet potato tubers or banana fruit) are combined
with leaves which normally have well balanced arrays of amino
acids, it is possible to reduce the overall level of protein in the
diet since the amino acid array in such a combination of feeds will
more closely approximate to that in the "ideal" protein (See Wang
and Fuller 1989).
Unfortunately much of the research with protein-rich forages has
been with conventional energy sources (maize and rice bran). As
these feeds contain fibre and are imbalanced in essential amino
acids, the potential to use high levels of protein-rich foliages is
compromised. It is strongly recommended that future research in
this area should be with basal diets that are low in both protein
and in cell wall constituents, as this will facilitate: (i)
increasing the percentage of forage protein in the diet; and (ii)
decreasing the overall protein level in the diet in accordance with
the concept of the "ideal" protein.
There is also a need to establish more accurately the levels of
nutrients in protein-rich leaves, especially the proportions of the
most limiting essential amino acids. An attempt to summarize some
of these data (Table 1) revealed a great deal of variation with
contradictory values being reported by authors even within the same
publication. Part of the problem appears to be the confusion when
authors report only levels of methionine, which may or may not also
include values for cystine.
Table 1: Major essential AA in the ideal
protein, soybean meal and leaves of selected protein-rich leaves
Ideal protein(1)
Soybean meal (2)
Water spinach (3)
Cassava leaves (4)
Sweet potato leaves (5)
Duckweed (7)
New cocoyam (6)
Mulberry
(3)
g AA/kg N*6.25
Lysine
63.2
42.7
56-65
39
43
46
50.6
Methionine
13.5
18-21
16.3
27.9
14.3
16.5
Cystine
10.3
15-16
5.27
7.38
12.6
12.0
Met+Cys
28.3
23.8
33-37
39
35.3
26.9
28.6
Threonine
38.9
39.5
47
51
42
39.5
45.1
As proportion of
lysine = 100
Lysine
100
100
100
100
100
100
100
100
Met+Cys
59
44.8
56
53-57
55
82
58.5
56.4
Threonine
75
61.6
92
76
114
98
85.6
89.1
Composition of
fresh leaves, g/kg fresh matter
DM
83
320
161
62
180
261
Composition, g/kg
DM
Crude protein
51.8
267
245
282
370
248
222
Crude fibre
31
155
156
128
77
142
172
Ash
62
142
84
109
16
133
126
(1)Wang and
Fuller 1989; (2) Martin 1990 ; (3) Phiny et al 2003; Phiny 2006,
personal communication; (4) Eggum 1970; (5) Woolfe 1992; (6)Rodríguez et
al 2006; (7) Le Thi Men 2006
What appears to categorise the most
useful sources of leaf protein, as identified by animal trials in
this review, are a relatively low level of crude fibre and a ratio
of sulphur amino acids relative to lysine close to that in the
ideal protein. The wide differences found in many reports for
amino acid levels in cassava leaves and sweet potato leaves (the
most reliable values for cassava leaves were done 36 years ago)
emphasize the need for a coordinated research effort in which
common samples of the most useful protein-rich leaves are
distributed for analysis to several laboratories where the
necessary equipment and expertise are available.
References
An L V 2004 Sweet potato leaves for growing pigs: Biomass
yield, digestion and nutritive value. Doctor's dissertation. ISSN
1401-6249, ISBN 91-576-6750-0
Berg C 2001 World Ethanol Production Fo Licht's
International Molasses and Alcohol Report . http://www.fo-licht.com
Bui Huy Nhu Phuc, Ogle R B, Lindberg J E and Preston T R
1996 The nutritive value of sun-dried and ensiled cassava
leaves for growing pigs. Livestock Research for Rural
Development Retrieved (8) 3, Retrieved from http://www.cipav.org.co/lrrd/lrrd8/3/phuc83.htm
Chhay Ty and Preston T R 2005aEffect of water spinach and
fresh cassava leaves on intake, digestibility and N retention in
growing pigs. Livestock Research for Rural Development. Vol.
17, Art. #23. Retrieved June 30, 2005, from http://www.cipav.org.co/lrrd/lrrd17/2/chha17023.htm
Chhay Ty and Preston T R 2005b: Effect of water spinach
and fresh cassava leaves on growth performance of pigs fed a basal
diet of broken rice. Livestock Research for Rural Development.
Volume (17) 7. Retrieved , from http://www.cipav.org.co/lrrd/lrrd17/7/chha17076.htm
Du Thanh Hang 1998 Ensiled cassava leaves and duckweed as
protein sources for fattening pigs on farms in Central Vietnam.
Livestock Research for Rural Development. Volume (10) 3.
Retrieved , from http://www.cipav.org.co/lrrd/lrrd10/3/hang2.htm
Du Thanh Hang and Preston T R 2005The effects of simple
processing methods of cassava leaves on HCN content and intake by
growing pigs. Proceedings of Workshop-seminar "Making better useof
local feed resources" MEKARN-CTU, Cantho, 23-25 May, 2005. http://www.mekarn.org/procctu/hang.htm
Eggum B O 1970 The protein quality of cassava leaves.
British Journal of Nutrition, Volume 24, Page 761
Ishida H, Suzano H, Sugiyama N, Innami S, Tadokoro T and
Meakawa A 2000 Nutritive value on chemical components of
leaves, stalks and stems of sweet potato (Ipomoea batatas
Poir). Food Chemistry 68: 359-367.
Kean Sophea and Preston T R 2001 Comparison of
biodigester effluent and urea as fertilizer for water spinach
vegetable. Livestock Research for Rural Development 13 (6) http://www.cipav.org.co/lrrd/lrrd13/6/kean136.htm
Le Thi Men 2006 Genetic and nutritional diversity of
animal and plant resources as animal feed and their utilization in
swine husbandry in the Mekong Delta of Vietnam. Doctoral thesis.
Tokyo University of Agriculture and Technology, Japan.
NRC 1988 Nutrient Requirements of Swine (9th Ed.).
National Academy Press, Washington, DC.
Pearse Lyons T and Bannerman J 2001The US Fuel Alcohol
Industry from 1980 to 2001; Lessons for other markets . In: A time
for Answers. Proceedings of Alltech's 15th Asia-Pacific
Lecture Tour 115\
Phiny C, Preston T R and Ly J 2003 Mulberry (Morus
alba) leaves as protein source for young pigs fed rice-based
diets: Digestibility studies; Livestock Research for Rural
Development (15) 1. Retrieved , from http://www.cipav.org.co/lrrd/lrrd15/1/phin151.htm
Phuc B H N, Lindberg J E, Ogle B and Thomke S
2001 Determination of the nutritive value of biomass products
for monogastrics using rats. Part 1. Comparison of eight forage
species at two level of inclusion in relation to a casein diet.
Asian Australasian Journal of Animal Science. 14, 896-993
Pimental D and Patzek T W 2005 Ethanol production using
corn, switchgrass, and wood; biodiesel production using soybean and
sunflower. Natural Resources Research 14(1):65-76.
PrestonT R 1989 Food, fuel and jobs from sugar
cane and tree legumes. In: New Crops for Food and Industry (edited
by G E Wickens, N Haq, P Day) 1989, 444 p. Springer Publishers.
PrestonT R and Rodríguez Lylian 2004:
Production and utilization of cassava foliage for livestock in
integrated farming systems. Livestock Research for Rural
Development. Vol. 16, Art. #28. Retrieved , from http://www.cipav.org.co/lrrd/lrrd16/5/pres16028.htm
Rameker J 2004 Grain production, supply and demand
outlook and trends in livestock feed industry. Proceedings 11th
Animal Science Congress, Kuala Lumpur, Malaysia, September 5-8,
2004
Rodríguez Lylian and Preston T R 1996 Comparative
parameters of digestion and N metabolism in Mong Cai and Mong
Cai*Large White cross piglets having free access to sugar cane
juice and duck weed. Livestock Research for Rural Development
(8) 1. Retrieved , from http://www.cipav.org.co/lrrd/lrrd8/1/lylian.htm
\
Rodríguez Lylian, Lopez D J, Preston T R and Peters K
2006 New Cocoyam (Xanthosoma sagittifolium) leaves as
partial replacement for soya bean meal in sugar cane juice diets
for growing pigs. Livestock Research for Rural Development.
Volume 18, Article #91. Retrieved September 11, 2006, from http://www.cipav.org.co/lrrd/lrrd18/7/rodr18091.htm
Speer V C 1990
Partitioning nitrogen and amino acids for
pregnancy and lactation in swine: A review. Journal of Animal
Science 68: 553-561.
Wang T C and Fuller M F 1989 The optimum dietary amino acid
pattern for growing pigs. British Journal of Nutrition. 62,
17-89
Woolfe J A 1992 Sweet potato: an untapped food resource.
Cambridge University Press, Cambridge, 643 pp.