|
|
|
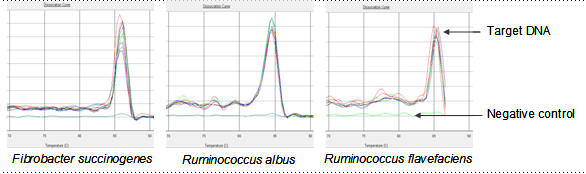
Figure 1: Dissociation curves showing specificity of real-time PCR in quantifying cellulolytic bacteria in the rumen of cattle |
| Back to contents |
Defaunation, the removal of protozoa from the rumen was used to investigate the effects of protozoa on rumen bacteria. Twelve Sindhi x Yellow cattle were allowcated into a two by two factorial design (absence or presence of protozoa; cottonseed cake or concentrate supplement). The basal rougahe diet was based on natural grass and rice straw. The animals were adapted for 21 days with the experimental diets before being dosed with soybean oil, a defaunation agent, at 6 ml/kg live weight. Rumen fluid was collected at day 5, 30,60 and 90 post defaunation. Conventional counting of protozoa was performed. In additon, genomic DNA rumen microorganisms was extracted to further used in real-time PCR quantification of cellulolytic bacteria.
Protozoa numbers significantly reduced at 5 days after oil drench and gradually recovered until day 90. Relative fold changes of Ruminococcus albus and Ruminococcus flavefaciens were significanly higher in defaunated cattle at both protein source supplements but the number of Fibrobacter succinogenes was lower (0.84-0.39 fold) in defaunated cattle under concentrate supplementation. Moreover, defaunation did not show significant effects on the abundance of methanogens, with an exception of increasing fold change at day 5 in defaunated cattle.
It is concluded that partial defaunation using soybean oil was beneficial regarding to an increasing numbers of R. albus and R. flavefaciens but the advantagous evidence on methanogens abundance was not achieved.
Rumen microorganisms comprise a diverse population of anaerobic bacteria, ciliate protozoa, fungi and archaea that account for most of the fermentative activities in the rumen to supply nutrients for the ruminant (Hungate 1966). In this microbial system, the interaction among the microbes is of importance as it will affect the efficiency of feed use and production of microbial end products available for the host animal. Of these microbes, protozoa constitute a large proportion of biomass, up to 40% in some animals (Theodorou and Frank 2005) especially their presence (faunated) or absence (defaunated) may influence on the abundance of certain baterial species and fungi. For instance, Takenaka and Itabashi (1995) reported that elimination of protozoa result in an increase of total bacteria but a decrease in methanogenic archaea. In an in sacco experiment it was found that defaunation increase activity of cellulolytic bacteria (Hobson 1988) and this was later confirmed by Arakaki et al (1995), who found a remarkable reduction of amylolytic and cellulolytic bacteria when unfaunated cattle are inoculated with protozoa.
In addition to being investigated in rumen function, protozoa has been studied on their effect on methane production. Protozoa has been shown to account for 9-25% of methanogenesis in the rumen fluid (Newbold et al 1995) and thus their absence from the rumen may contribute to methane production to the environement (Ushida and Jouany 1996). However, it could be noted that the magnitude of methane reduction or the efficacy on cattle from elimination of protozoa is diet-dependent and this technique is appropriate for a short feeding period, i.e. in fattening stage since it is difficult to maintain defaunated animals, especially under household level – where cattle are mainly fed on poor quality feeds. In this case, the combination of feeding stratergy such as including cottonseed cake, a source of bypass protein or concentrate in the diet and defaunation technique may be a good option.
With the development of molecular techniques like real-time PCR, the study on functional microbes without cultivation has become easier and more convenient. One of the first demonstrations of the applicability of real-time PCR for quantification of bacteria species was by Tajima et al (2001), in which they could monitor the corresponding bacteria species in cow rumen when the diet was switched from hay to grain. Similarly, Koike and Kobayashi (2001) successfully enumerated three cultured rumen cellulolytic species Fibrobacter succinogenes, Ruminococcus albus and Ruminococcus flavefaciens. Additionally, Denman and McSweeney (2006) developed a real-time PCR assay to monitor the fungal and cellulolytic bacteria populations in the rumen of Brahman crossbreed steers. Using this technique, the aim of the present study was to understand the shift or fold changes of rumen cellulolytic bacteria and methanogens over total bacteria in cattle with either cottonseed cake or concentrate supplementation in faunated and defaunated condition.
Twelve crossbred Sindhi cattle weighing from 140 – 160 kg were selected and arranged in two by two factorial design (absence and presence of protozoa; with either cottonseed cake of concentrate supplement). Each treatment includes 3 animals and the experimental layout is as follows:
Treatment 1: basal diet + 0.5% cottonseed cake + partial defaunation
Treatment 2: basal diet + 0.5% cottonseed cake + undefaunation
Treatment 3: basal diet + 0.5% concentrate + partial defaunation
Treatment 4: basal diet + 0.5% concentrate + undefaunation
The partical defaunation factor was soybean oil with a single dose of 6 ml/kg live weight at the beginning of the experiment. The supplementation of either cottonseed cake or concentrate was calculated on live weight basis. Basal diet was natural grass, which was offered twice a day adjusted to intake during the adaptation period. Rice straw was freely accessed in the evening. Concentrate composition included 42% rice bran, 10% corn starch, 25% tapioca, 10% soy, 10% mollases, 2% urea and 1% salt, which finally offered the nutrient values of about 83% dry matter, 14% crude protein and 35% neutral detergent fiber. During the experiment, the animals were totally fed in individual pen, drinking water and home-made mineral blocks were available all the time.
The animals were kept 21 days for adaptation period followed by a soybean oil drench (defaunation) at 6 ml/kg live weight. Rumen samples were collected at 5, 30, 60 and 90 days after defaunation. Rumen fluid (about 100 ml) were obtained before feeding by using a suction pump and a stomach tube with a brass filter. Samples were mixed with an equal volume of 2% formalin in phosphate-buffer saline for conventional enumeration using microscopy and the other aliqout were immediately kept in 70% ethanol for further DNA extraction.
Microscopic countings of protozoa: protozoa biomass were determined by staining two ml of rumen fluid with methylene blue and counting in a Malasser cell (0.2 mm chamber) under a microscope at 10x magnification.
DNA extraction from rumen fluid: genomic DNA extraction was done following the CTAB-based protocol described by Minas et al (2011). In brief, the cell lysis was achieved by bead beating in the presence of CTAB lysis buffer. Phase was separated by centrifugation with the presence of phenol: chloroform: ioamyl alcohol (25: 24: 1, pH = 6.7; Sigma-Aldrich). Acid nucluic was further precipitated by adding isopropanol containing 50 µl of 7.5 M ammonium acetate (Fisher). DNA pellet was washed by ethanol and air-dried and resuspended in 200 µl of 75 mM TE buffer.
Primers and real-time PCR: species-specific primers and PCR conditions used to amplify partial 16S rDNA regions were picked up from liturature (Table 1). Real-time PCR was performed on an ABI Prism 7000 Sequence Detection System (Applied Biosystems). All experiments were done in duplicate. The fold difference of certain bacteria species was analyzed by the the two ddCt method (Livak and Schmittgen 2001) and the data were calculated as fold change relative to the control (faunated cattle), which was arbitrarily assigned as 1.
The effects of treatment on rumen microorganisms were subjected to unpaired 2-tailed Student’s t-test or one way ANOVA in Minatab 13.2 where appropriated.
|
Table 1: Real-time PCR primers and conditions for amplification of rumen bacteria |
|||
|
Target bacterium |
Primer sequence |
Tm, Real-time PCR product |
References |
|
Total bacteria |
Fw: CGGCAACGAGCGCAACCC Rev: CCATTGTAGCACGTGTGTAGCC |
60oC, 130 bp |
Denman and McSweeney (2006) |
|
Fibrobacter succinogenes |
Fw : GGTATGGGATGAGCTTGC Rev: GCCTGCCCCTGAACTATC |
60oC, 446 bp |
Koike and Kobayashi (2001) |
|
Ruminococcus albus |
Fw: CCCTAAAAGCAGTCTTAGTTCG Rev: CCTCCTTGCGGTTAGAACA |
60oC, 175 bp |
Koike and Kobayashi (2001) |
|
Ruminococcus flavefaciens |
Fw: TCTGGAAACGGATGGTA Rev: CCTTTAAGACAGGAGTTTACAA |
55oC, 295 bp |
Koike and Kobayashi (2001) |
|
Methanogens |
Fw: TTCGGTGGATCDCARAGRGC Rev: GBARGTCGWAWCCGTAGAATCC |
60oC, 140 bp |
Denman et al (2007) |
|
Fw: Forward primer; Rev: Reverse primer |
|||
Figure 1 shows the dissociation curves of specific primers in amplification of three cellulolytic bacteria namely Fibrobacter succinogenes, Ruminococcus albus and Ruminococcus flavefaciens. These curves were in single peak and confirmed the specificity of primers in quantification of bacteria species reported by Koike and Kobayashi (2001).
|
|
|
Figure 1: Dissociation curves showing specificity of real-time PCR in quantifying cellulolytic bacteria in the rumen of cattle |
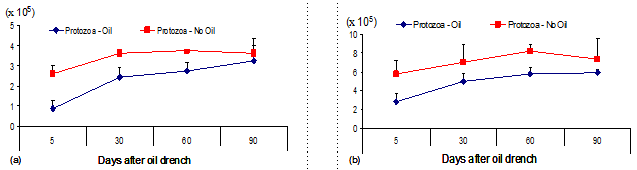
In our previous studies, it was shown that drenching cattle with 6 ml oil/kg live weight was appropriate for eliminating the protozoa and resulted in better performance of fattening cattle; although this method did not resulted in complete defaunation, the benefit was obtained over the fattening period of 90 days (Nguyen Thi Hong Nhan et al 2007; 2008). In this report, similar outputs were achieved, of which protozoa numbers were significantly reduced at day 5 after oil treatment and gradually recovered until day 90 (Table 2; Figure 2). It was additionally noted that under both faunated and partial defaunation condition, cattle supplemented with concentrate did show a trend of being higher protozoa numbers than those received cottonseed cake in the diet.
|
Table 2: Microscopic count of protozoa numbers in faunated and partially defaunated cattle |
||||
|
Treatment |
Days afer oil drench |
|||
|
5 |
30 |
60 |
90 |
|
|
Cottonseed cake supplemention |
|
|
|
|
|
Oil |
0.87 ± 0.41 |
2.43 ± 0.47 |
2.73 ± 0.40 |
3.23 ± 0.76 |
|
No oil |
2.60 ± 0.60 |
3.60 ± 0.53 |
3.77 ± 0.40 |
3.60 ± 1.74 |
|
P |
0.015 |
0.046 |
0.035 |
0.754 |
|
Concentrate supplemention |
|
|
|
|
|
Oil |
2.85 ± 0.87 |
5.00 ± 0.87 |
5.80 ± 0.70 |
5.93 ± 0.32 |
|
No oil |
5.80 ± 1.42 |
7.07 ± 1.90 |
8.23 ± 0.65 |
7.40 ± 2.21 |
|
P |
0.037 |
0.161 |
0.012 |
0.319 |
|
|
|
Figure 2: Changes of protozoal numbers over the 90-day period in partially defaunated and faunated cattle supplemented with (a) cottonseed cake and (b) concentrate in the diet |
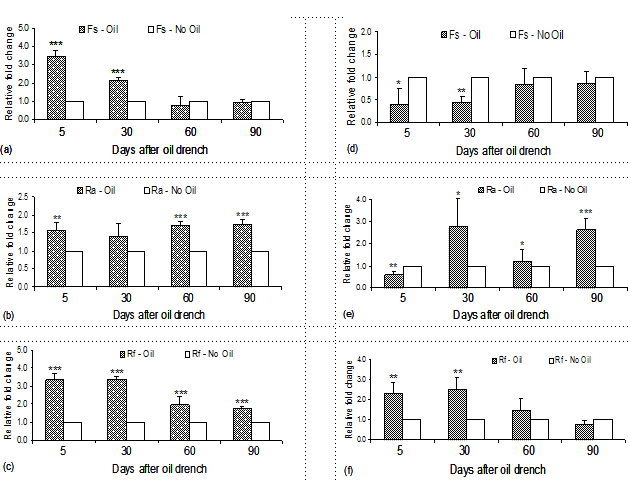
The shift of cellulolytic bacteria in cattle under faunated and partial defaunated condition and different protein source supplements is presented in Table 3 and Figure 3. Both oil and supplement factors appeared to have certain effects on the population of rumen bacteria. For F. succinogenes, adverse shift of bacteria number was obtained at day 5 and day 30 after oil drench, in which cottonseed cake led to an increase of Fibrobacter succinogenes whereas a significant decrease was observed in cattle supplemented with concentrate. However, these changes did not exist at later stages (day 60 and 90) where the fold discrimination between two cattle groups was almost similar and close to 1. In contrast, both R. albus and R. flavefaciens were beneficial regarding to their abundance in the rumen, but greater fold change stability was examined in cattle given cottonseed cake.
|
Table 3: Relative changes of rumen bacteria by real-time PCR |
|||||||
|
Days after oil drench |
Relative fold change1 |
SEM |
P |
|
Relative fold change2 |
SEM |
P |
|
|
Fibrobacter succinogenes |
|
Fibrobacter succinogenes |
||||
|
5 |
3.42 |
0.214 |
0.000 |
|
0.39 |
0.196 |
0.036 |
|
30 |
2.13 |
0.084 |
0.000 |
|
0.43 |
0.09 |
0.003 |
|
60 |
0.76 |
0.267 |
0.414 |
|
0.82 |
0.205 |
0.437 |
|
90 |
0.94 |
0.075 |
0.447 |
|
0.84 |
0.163 |
0.392 |
|
|
Ruminococcus albus |
|
Ruminococcus albus |
||||
|
5 |
1.57 |
0.119 |
0.009 |
|
0.60 |
0.084 |
0.009 |
|
30 |
1.41 |
0.206 |
0.117 |
|
2.74 |
0.644 |
0.036 |
|
60 |
1.71 |
0.075 |
0.000 |
|
1.21 |
0.301 |
0.523 |
|
90 |
1.74 |
0.072 |
0.000 |
|
2.62 |
0.242 |
0.001 |
|
|
Ruminococcus flavefaciens |
|
Ruminococcus flavefaciens |
||||
|
5 |
3.35 |
0.571 |
0.015 |
|
2.28 |
0.312 |
0.015 |
|
30 |
3.39 |
0.397 |
0.004 |
|
2.50 |
0.299 |
0.002 |
|
60 |
1.95 |
0.214 |
0.011 |
|
1.46 |
0.295 |
0.174 |
|
90 |
1.74 |
0.009 |
0.000 |
|
0.76 |
0.091 |
0.038 |
|
1, 2 Cattle supplemented with cottonseed cake and concentrate, respectively |
|||||||
In an earlier study by Mosoni et al (2011), it was clearly shown that long- and short-term defaunation increased nearly 1 log of the total rumen bacteria in sheep. In this study, such an effect was not observed over the 90-day period (data not shown), but defaunation has resulted in a modification of cellulolytic community abundance. In fact, many reports have mentioned on an increase of cellulolytic bacteria in defaunated ruminants (Ushida et al 1991; Arakaki et al 1995). In the present work, this bacteria community appeared to follow that trend, especially in cattle supplemented with cottonseed cake, of which three bacteria species examined did show a higher fold change between the defaunated and faunated cattle groups. On the other hand, while the number of R. albus and R. avefaciens increased a remarked reduction of F. succinogenes was detected in rumen of cattle consumed concentrate. This finding supported the work of Monosi et al (2011) that defaunation increased the content of R. albus and R. flavefaciens up to 2 log (similar to 2 fold changes) but did not affect on F. succinogenes.
|
|
|
Figure 3: Relative fold change (oil/no oil) of cellulolytic bacteria numbers namely Fibrobacter succinogenes, Ruminococcus albus and Ruminococcus flavefaciens, respectively over the 90-day period in cattle supplemented with cottonseed cake (a, b and c) and concentrate (d, e and f) |
|
*** P < 0.001; ** P < 0.1; * P < 0.05 |
Previously, Ozutsumi et al (2006) also reported similar shift of direction in sheep, where greater gram-positive bacteria (R. albus and R. flavefaciens) but lower gram-negative bacteria (F. succinogenes) were available in defaunated sheep. The reason is not clear but these authors concluded that defaunation did not always bring benefits of increasing the population of cellulolytic bacteria. Additionally, in sheep fed a high concentrate diet, no differences found on F. Succinogenes and other cellulolytic bacteria numbers between faunated and protozoa-free group (Yáñez-Ruiz et al 2009). Thus, it is evidenced that the abundance of these major cellulolytic bacteria was differentially affected by the absence of protozoa and it could be that another cellulolytic species present in domesticated ruminants as there has been a report on a new and very active cellulotyic bacteria available in yak (Cai et al 2010).
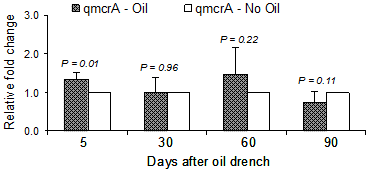
A significant effect of defaunation on numbers of methanogens was found at day 5, where there was an evidence of increasing methanogens in the rumen (Figure 4). At other time points of rumen collection, the fold change between faunated and partially defaunated cattle remained similar. This was somewhat unexpected since it was widely accepted that in the absence of protozoa, methane emissions reduced up to 20% (Morgavi et al 2008). The present results are in agreement with those of Mosoni et al (2011) on the fact that methanogens abundance increased approximately 1 log in defaunated sheep; however in that study a stable reduction of methane production was observed. From these findings, these authors and others (Firkins and Yu 2006) suggested that the reduction of methane emissions has not been necessarily associated with a lower amount of methanogens in the rumen. Further work need to be done to figure out the relationship between these two factors under different feeding and management conditions.
|
|
|
Figure 4: Relative fold change of methanogens in cattle on cottonseed cake supplementation |
Based on the results of this research it is concluded that:
· Abundance of cellulolytic bacteria increased in partial defaunated cattle with an exception of F. succinogenes in cattle under concentrate supplementation.
· There was an increase of methanogens in defaunated cattle at 5 days after oil drench, otherwise it remained similar at both faunated and defaunated condition.
The authors are grateful to the MEKARN project financed by the Sida-SAREC agency and the project B2011-16-05 funded by the Ministry of Education and Training of Vietnam.
Arakaki C, Mitsumori H, Itabashi H, Shirasaka S and Minato H 1995 Influence of the presence of protozoa on the rumen microbial population of cattle. Journal of General and Applied Microbiology 40: 215-226.
Cai S, Li J B, Hu F Z, Zhang K G, Luo Y M, Janto B, Boissy R, Ehrlich G and Dong X Z 2010 Cellulosilyticum ruminicola, a newly described rumen bacterium that possesses redundant fibrolytic-protein-encoding genes and degrades ligno-cellulose with multiple carbohydrate-borne fibrolytic enzymes. Applied and Environmental Microbiology 76: 3818-3824.
Denman S E, Tomkins N W and McSweeney C S 2007 Quantitation and diversityanalysis of ruminal methanogenic populations in response to the anti-methanogenic compound bromochloromethane. FEMS Microbiology Letters 62: 313–322.
Firkins J L and Yu Z 2006 Characterisation and quantification of the microbial populations in the rumen. In Ruminant Physiology, Digestion, Metabolism and Impact of Nutrition on Gene Expression, Immunology and Stress. Sejrsen K, Hvelplund T and Nielsen M O, ed. Wageningen Academic Publishers, Wageningen, the Netherlands. Pp 19-54.
Hobson P N and Jouany J P 1988 Models, mathematical and biological, of the rumen function. In: The Rumen Microbial Ecosystem. Edited by Hobson P N, Elsevier Applied Science, London pp. 461-511.
Hungate R E 1966 The rumen protozoa. In “The Rumen and Its Microbes”, Academic Press, New York, pp. 91-147.
Livak K J and Schmittgen T D 2001 Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods 25(4): 402-408.
Nguyen Thi Hong Nhan, Nguyen Trong Ngu, Nguyen Thiet, Preston T R and Leng R A 2007 Determination of the optimum level of a soybean oil drench with respect to the rumen ecosystem, feed intake and digestibility in cattle. Livestock Research for Rural Development. Volume 19, Article #117. Retrieved April 25, 2011, from http://www.lrrd.org/lrrd19/8/nhan19117.htm.
Nguyen Thi Hong Nhan, Nguyen Trong Ngu, Preston T R and Leng R A 2008 Effects of drenching soybean oil and fish oil on intake, digestibility and performance of cattle fattening in the Mekong Delta, Vietnam. Volume 20, Article #113. Retrieved April 25, 2011, from http://www.lrrd.org/lrrd20/7/nhan20113.htm
Minas K, McEwan N R, Newbold C J, Scott K P 2011 Optimization of a high-throughput CTAB-based protocol for the extraction of qPCR-grade DNA from rumen fluid, plant and bacterial pure cultures. FEMS Microbiology Letters 325(2):162-9.
Morgavi D P, Jouany J P and Martin C 2008 Changes in meth-ane emission and rumen fermentation parameters induced by refaunation in sheep. Australian Journal of Experimental Agriculture 48: 69–72.
Mosoni P, Martin C, Forano E and Morgavi D P 2011 Long-term defaunation increases the abundance of cellulolytic ruminococci and methanogens but does not affect the bacterial and methanogen diversity in the rumen of sheep. Journal of Animal Science 89: 783-791.
Newbold C J, Lassablas B and Jouany J P 1995 The importance of methanogens associated with ciliate protozoa in ruminal methane production in vitro. Letters in Appllied Microbiology 21: 230-234.
Ozutsumi Y, Tajima K, Takenaka A and Itabashi H 2006 Real-time PCR detection of the effects of protozoa on rumen bacte-ria in cattle. Current Microbiology 52: 158-162.
Tajima K, Aminov R I, Nagamine T, Matsui H, Nakamura M and Benno Y 2001 Diet-dependent shifts in the bacterial population of the rumen revealed with real-time PCR. Applied and Environmental Microbiology 67:2766–2774.
Takenaka A and Itabashi H 1995 Changes in the population of some functional groups of rumen bacteria including methanogenic bacteria by changing the rumen ciliates in calves. Journal of General and Applied Microbiology 41(5): 377-387.
Theodorou M K and Frank J 2005 Rumen micro-organisms and their interactions. In: Quantitative Aspects of Ruminant Digestion and Metabolism. Edited by Dijkstra J, Forbes J M, France J, 2nd edn: CABI Publishing; 2005: 736.
Ushida K and Jouany J P 1996 Methane production associated with rumen-ciliated protozoa and its effect on protozoan activity. Letters in Applied Microbiology 23: 129-132.
Ushida K, Jouany J P and Demeyer D I 1991 Effects of presence or absence of rumen protozoa on the efficiency of utilization of concentrate and fibrous feeds. In Physiological Aspects of Digestion and Metabolism
in Ruminants: Proceedings of the 7th International Symposium on Ruminant Physiology, pp. 625-654 (Tsuda T, Sasaki Y and Kawashima R, editors) San Diego: Academic Press.
Yanez-Ruiz D R, Martin-Garcia A I, Hart K J, Molina-Alcaide E and Newbold C J 2009 Cellulolytic activity in the rumen of lambs fed a high concentrate diet is not affected by the removal of protozoa. In: Papachristou T G; Parissi Z M; Ben Salem H; Morqand-Fehr P (Eds). Options Mediterraneenes, Serie A, No. 85, Nutritional and foraging ecology of sheep and goats, pp. 251-254.