 |
|
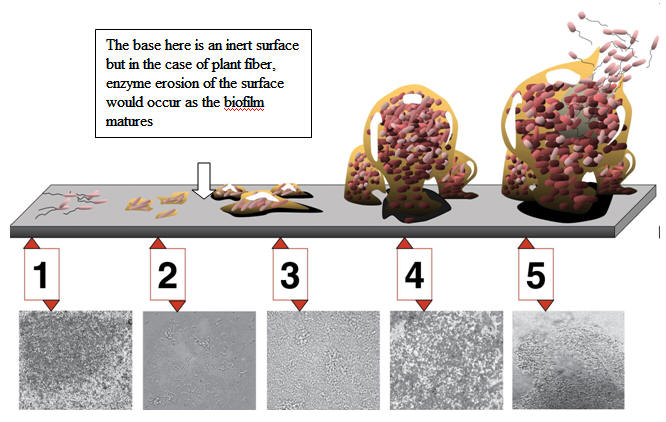
Figure 1. (Anonymous 2011) http://microbialecologynotes.wordpress.com/tag/microbial-ecology/ |
The rumen has been viewed as a fermentation vat with free floating (planktonic) microbes, particle associated microbes and microbes that attach to the epithelial cells of the rumen wall. The rumen is described, in general, as a, anaerobic microbial ecosystem containing a milieu of bacteria, archaea, protozoa, bacteriophage and phycomycetous fungi that degrade organic matter with the ultimate production of volatile fatty acids (VFA),the energy that becomes available is used for maintenance and growth of cells. The VFA are absorbed from the upper part of the digestive tract and the microbial cells provide essential nutrients such as amino acids, when they are digested by intestinal enzymes on wash out from the rumen. Since about 1980 there has been an emerging realization that possibly as high as 80-90% of the rumen microbial biomass is present as associated and interactive colonies in biofilms on both inert and insoluble organic matter in the rumen and that the planktonic cells play little role in feed digestion.
Biofilms consist of microbial cells and a wide range of self-generated extra- cellular polymeric substances. Biofilm formation is a dynamic process, which is coordinated by the interactions of different microbial species. These biofilms that form within a few minutes of feed being immersed in rumen fluid, provide spatially separated microbial colonies held in an extracellular polymeric substance (EPS), including polysaccharides, nucleic acids, and proteins that is progressively produced by the microbes as they position themselves in the biofilm. The sequential deposition and growth of sessile microbes within the biofilm matrix allows the nutrients produced as end products of one species to be efficiently transferred to other species which use these as substrate for energy metabolism. The overall conversion of plant organic matter to VFA in the biofilm results in the production of reduced cofactors which revert to their oxidized form by releasing the electron as hydrogen. This depends on the maintenance of a low concentration of hydrogen and requires rapid uptake of hydrogen by microbes that can use high affinity electron acceptors such as nitrate, sulphate and carbon dioxide. Methanogenic Archae which obtain energy by reducing carbon dioxide are mainly responsible, but sulphur reducing bacteria also play a minor role in the rumen. The requirements for minimal concentrations of hydrogen in the culture medium demand that the syntrophic microbes are closely associated. Syntrophism is a special case of symbiotic cooperation between two metabolically different types of bacteria which depend on each other for degradation of a certain substrate, typically for energetic reasons. Biofilm formation on feed particulate matter is essential for efficient fermentative digestion in the rumen, as it provides the ecological niche that ensures that the synrophic organisms involved are spatially aligned to ensure efficient capture of hydrogen to prevent feed back inhibition of the reactions involved in converting polymeric feed materials to their end products.
The overall conclusion pertinent to this symposium is that because of the need for symbiotic associations in biofilm microbial colonies on feed particles in the rumen, it is not logical to attempt to lower methane production by including chemicals in a diet that specifically target the inhibition of methanogens, unless alternative electron sinks are provided. Where compounds that can be used by microbes as electron sinks are included in the culture media , the organism using these as sources of energy must also be spatially distributed in the biofilm to maintain hydrogen concentrations at a minimum for the efficient oxidation of reduced cofactors generated in the reactions occurring in fermentation. Nitrate reducing and sulphur reducing bacteria appear to full fill this role highly efficiently.
The contents of the rumen have been historically viewed as a milieu or microbial soup which collectively is very effective in efficiently degrading the many feed resources to VFA with the growth of microbial cells. There has been a gradual change in this perception as the concept of microbial communities in organized and structured systems have become recognized for the control of the complex hydrolytic breakdown of feed particles (see Costerton et al 1987).
Ruminant nutritionist’s perception of rumen bacteria as unicellular life forms is deeply rooted in the pure-culture paradigm. Rumen fluid can be diluted and studied in liquid culture; this mode of operation has been exploited and used to study bacteria in terms of their substrate and metabolic end products. This traditional way of culturing bacteria in liquid medium or on agar plates has been instrumental in allowing the study of microbial fermentation and ha been enlightening as to the microbes that are involved and the pathways by which these microbes convert materials, indigestible in the monogastric animal, to nutrients that are used for energy metabolism and synthesis of tissues in ruminant animals
Planktonic growth of bacteria is, however, rarely how most bacteria exist in nature (see Nikolaev and Plakunov 2007) and in all probability how they function in the rumen (see Edwards et al 2008). Environmental microbiologists have long recognized that complex bacterial communities are responsible for driving all the major nutrient cycles within the earth’s biosphere including carbon, sulphur and nitrogen biogeochemical cycling that maintains the biosphere (see Davey and O’Toole 2000). Until recently, the lack of methods for exploring these communities in situ has hampered detailed analyses. Application of technologies independent of having to culture the microbes has illustrated that the variety of bacterial species present in the rumen has been vastly underestimated (Rappe and Giovanni 2003; see Edwards et al 2008).
Over the last 50 years rumen microbiologists have emphasized the need for attachment of bacteria to feed materials in the efficient digestion of feed. Measurements of the sites of bacterial ATP and its location in the rumen indicated that the majority of this was associated with plant particles (Forsberg and Lam 1977;Craig et al 1987) and isotope dilution studies using organisms labeled with 15Nsuggests that 80-90 % of the bacteria are associated with particulate matter (Rodriguez et al 2003). Krebs (1987) labeled bacteria with 35S-sulphate and examined the movement of 35S between the free floating and particle associated organisms. This research has been recently re-examined using a modeling approach by Nolan (2011) who found evidence that indicated that 80% of the microbes washed out of the rumen originated from particle associated microbes and that bacteria freely exchange between particles in the rumen without entering the free floating bacterial pool ( Krebs 1987). It is now clear that solid associated microbes play the most important role in digestion in the rumen (McAlister et al 1994 see also Edwards et al 2008) However the consequences of this have never been related to actual mechanisms of fermentative digestion that requires a number of different species of microbes to degrade the often complex structures of feed materials. Recently Wang and Chen (2009)and Weimer et al (2009) have contrasted the stark differences between the efficiencies of fermentative production of ethanol in the new biofuels industries based on cellulosic feed stock and the efficiency of the rumen organisms in converting cellulosic materials to the short chain VFA. Clearly the rumen has evolved highly efficient mechanisms for these processes (see Wang and Chen 2009).
Recent advances in microscopy and molecular technologies have made it possible to examine particle associated communities in situ in great detail and without the bias of liquid culture. Direct observation of a variety of natural anaerobic habitats including the rumen has established that the majority of microbes persist attached to surfaces within a structured biofilm ecosystem and not as free floating organisms (Mayorga et al 2007). These assemblages of bacteria in colonies within the biofilm matrix function as a cooperative consortium, in a complex way by providing substrate or removing end products that are inhibitory to one of the partners positioned in extracellular polymeric substances ( EPS) (see Costerton et al 1987;Costerton et al 1995a,b; Costerton 2007). Hence, although some rumen microorganisms may have an independent planktonic existence, there is growing evidence that they have an interdependent or syntrophic exisistance within the biofilm ( Cheng and Costerton 1980) in which they function as an integral part of the hydrolysis of polymers and conversion of feed materials eventually to VFA. Planktonic cells in rumen fluid seemingly being largely organism that breakaway from the biofilm mode of life because of declining substrate availability or possibly through erosion in rumen contractions or during eructation and rumination. This pool of organisms is then available for colonization and development of microbial biofilms on newly consumed feed particles or is washed into the lower tract with digesta movement.
The rumen is described in general as a microbial, anaerobic ecosystem containing a milieu of bacteria, archaea, protozoa, bacteriophage and phycomycetous fungi that form a multicultural community with each playing its part in the anaerobic fermentation of feed with the ultimate production of volatile fatty acids (VFA) via the Embden Meyerhof pathway. The continuous degradation of organic matter results in the production of reduced co-enzymes which if the concentrations of hydrogen are very low are oxidized with the release of electrons as hydrogen gas. This in turn is removed by methanogens bacteria that gain energy by reduction of carbon dioxide to methane. The VFA provides the major source of energy for metabolic processes in the animal. The energy (ATP) generated in fermenting organic matter being used for the maintenance and growth of microbial cells. The VFA are absorbed from the upper part of the digestive tract and the microbial cells provide essential nutrients such as amino acids, essential fatty acids and some vitamins when they are digested by intestinal enzymes on wash out from the rumen. Protein in forage is variably fermented to both VFA and branched chain VFA depending on the solubility of the protein source in plant tissues. Highly insoluble dietary protein in the particulate form may by-pass fermentation and provides substantial quantities of protein to the intestines for enzymatic hydrolysis to amino acids and absorption (see Preston and Leng 1986). Lipids are hydrolyzed to glycerol which is fermented and long chain fatty acids that are largely unchanged and sequester on to plant particles becoming available for absorption when these pass into the intestines.
More nutrients are extracted from residues of rumen fermentation and intestinal digestion by a further fermentative region in the caecum-large intestine before the feed, microbial residues, debris from the wall of the digestive tract and its unabsorbed secretions are excreted in the faeces.
From an ecological perspective, populations of bacteria arise from individual cells, and metabolically similar populations constitute groupings referred to as guilds (e.g.cellulolytic bacteria). Sets of guilds (e.g., cellulolytic and fermentative, sulfate- and sulfur-reducing, and methanogenic bacteria) conducting interdependent physiological processes form associated microbial communities. In essence, biofilms represent an interdependent community-based existence. Biofilms on the rumen wall or particulate matter in the rumen are derived from multiple microbial species, and they can form on a vast array of surfaces of inert or organic matter that is both living and dead. Microorganisms also form natural assemblages in suspensions, such as in sewage treatment works, anaerobic digesters and the rumen receiving liquid feeds, in which they preferentially aggregate to form flocs or granules (Macleod et al 1990:Rowe et al 1979).
For the purpose of this review, biofilms are broadly defined as assemblages of colonies of microorganisms and their associated extracellular products at an interface and typically attached to a surface of a particle that is only slowly solubilised or is inert The association of bacterial colonies within biofilm are essential for the syntrophic reactions that are required in a continuous fermentation process if end product inhibition or slowing of the rate of reactions are to be avoided and efficient and sufficiently rapid conversion of feed materials to nutrients are to be accomplished (see Stams and Plugge 2009).
Syntrophic association is one in which two or more organisms combine their metabolic capabilities to catabolize a substrate that cannot be catabolised by either one alone (Stams and Plugge 2009)
Whilst the rumen system has received limited work, in this presentation, the principles that apply widely in nature are presumed to apply in the rumen. Undoubtedly recent work with new sophisticated microscopy and tools for genetic identification of microbes (see Firkins 2010) have clearly shown that the biofilm mode of degradation of organic matter is dominant in the rumen (Edwards et al 2008). If this is correct then from published work and some extrapolations from the more researched biofilms of, for instance, sewage treatment systems can be justified that may increase our understanding of the rumen and how it can be manipulated to increase efficiency of feed utilization. In particular the biofilm environment is irreplaceable for efficient methanogenesis and this has major implications for mitigation of methane release in enteric fermentation. In this presentation information is extrapolated or drawn from research in related anaerobic ecosystems in particular the waste water treatment industries as well as that now published that reviews the rumen ecosystem (Edwards et al 2008).
Compounds such as cellulose with molecular weights in excess of 1000 atomic mass units are considered macromolecular and cannot be assimilated by bacteria without hydrolysis (Confer and Logan 1997). Thus cellulose digesting bacteria have to be the first colonizers of plant materials in the developing biofilm in the rumen. These organisms attach to feed particles where the waxy covering has been damaged and substrate is exposed and release enzymes to cleave the polymers of hemicellulose and cellulose and other macromolecules including proteins (see Costerton et al 1987) into shorter chain intermediates that are soluble ( sugars , peptides and amino acids). To make this process energy efficient substrate enzymes and microbes must remain within 500 micrometers of each other (Wetzel 1991). Physical contact between microbes and structural plant components is necessary in hydrolysis and cellulose degrading microbes attach to plant particulate matter and commence to produce extracellular polymeric substances or ECP. The ECP formed by these organisms develops into the biofilm matrix that provides the confined microenvironment keeping microbes, enzymes and substrates in close associations ( Fleming and Wingender 2001) enabling microbes within the biofilm to readily exchange substrates ( Minato et al 1966:Wolin et al 1997) and ensures a highly efficient environment for organic matter fermentation. For example cellulose breakdown when cellulolytic organisms are incubated in a suspended growth commercial digester (Hungate 1966 Chynoweth et al 2003) is only about 0.01 of that in rumen Digestion efficiency is improved almost 10 times when biofilm mode of utilization of cellulose is allowed in natural ecosystems The present evidence is indicative of the superiority of biofilm cellulose digestion over that achieved with planktonic organisms . In fact it would be impossible for ruminants to achieve the breakdown of sufficient plant materials to support life at the rates of utilization of cellulose recorded for suspended or planctonic cellulolytic microbes (see Wang and Chen 2009)
Under natural conditions, a solid material immersed in water containing organic matter is immediately covered by a primary (conditioning) microbial film, which provides a series of new properties to the surface (Davey and O’Toole 2008 ). The formation of this layer is the first stage of biofilm development and on inert surfaces is laid down prior to the attachment of cells and the formation of a bacterial film (Fig. 1). Where the surface is potentially digestible ( i.e. damaged plant tissue) cellulolytic organisms rapidly attach to the surface ( McAlister et al 1994).In the rumen, lingo-cellulosic material that have damaged surface layers or the surface where cellulose and other structural carbohydrates, sugars and protein are exposed are the primary sites of adhesion for the microbes that subsequently become the base layer of the biofilm (Cheng et al 1980).. However, it appears likely that biofilm may also form on the waxy un-reactive outer layer of leaf materials, particularly where a considerable proportion of a diet is highly soluble, such as with molasses based diets. Preston and Willis (1967) noted a requirement for fiber in molasses feeding systems which could be owing to the need for biofilm structures on particulate matter to efficiently ferment the soluble components of the feed
The next stage is reversible adhesion, when microorganisms become stuck with polymeric substances to the solid particle. The initial colonizer begins the process by synthesizing and excreting extracellular polymeric substances (EPS) which adhere to the anchored bacteria. Nonspecific physicochemical forces of interaction, act between the molecules and structures on the surfaces of the microorganism and the solid substrate (van derWaals’, hydrophobic, electrostatic, and London dispersion forces( Nikolaev and Plakunov 2007].In the rumen this may apply to the undamaged leaf material that is relatively inert but is secondary to the bacteria which attach to the inner digestible plant components that are exposed by damage to the surface layers during harvest or rumination and chewing of the plant materials by the animal.
The third phase consists of several independent stages of biofilm formation. In waste water systems for a certain period after attaching to the surface, the microbes can move along the surface by means of flagella and pili which themselves appear to interconnect the microbes (see Nikolaev and Plakunov 2007).This has not, however, been observed to occur in the rumen particulate biofilm. When the cells lose motility by attachment then some of them adhere to each other and they begin too produce and secrete extracellular polymer (polysaccharides, lipopolysaccharides, glycoproteins) [Sutherland 2001], which form the EPS, the basic material of the matrix of the biofilm.
The next colonizers are those that apparently use the products generated by the primary adhering microbes, i.e. microorganisms which locate in close association with the primary colonizers. Usually these organisms arrive and establish sessile colonies which appear to be organized and spatially arranged to sequentially use the products released into the EPS by the initial colonizers. These include sulphur reducing bacteria and nitrate reducing bacteria where these elements are in the bulk medium or released by primary colonizers from the substrate matrix of the organic matter of the particle. EPS production by the resident bacteria continues and forms specific structures: cavities, channels, outgrowths, and pores simultaneously with an increase in the thickness of the biofilm that traps and concentrates nutrients and enzymes and enables bacteria to readily exchange substrates. Under favorable conditions in, for instance, a water treatment rising sewer, this stage of the build-up of a mature biofilm continues and under unfavorable conditions, the biofilm enters its last stage, disintegration, degradation, and loss of some cells and liberation of the others ( see Figure 1).
In the rumen the dispersal or disruption of the biofilm may be brought about by the act of rumination and at the same time particle to particle exchange of biofilm fragments when the bolus is in the mouth, which may be instrumental in initial colonization of feed particles (see below).
The culmination of the development of the biofilm is when the bacteria become sessile (are fixed in a position in the matrix of the biofilm where the availability of substrate, cell growth and division increase) and closely.
|
|
Figure 1. (Anonymous 2011) http://microbialecologynotes.wordpress.com/tag/microbial-ecology/ |
1. Initial (reversible) attachment of cells to the surface by adhesins, receptors and non-specific mechanisms that rely on physical-chemical forces such as van-der Waals forces.
2. Irreversible attachment by production of EPS resulting in more firmly adhered.
3. Maturation I. Early development of biofilm architecture.
4. Maturation II. Maturation of biofilm architecture, attachment of other organisms, competition, organization to create pores, channels.
5. Dispersion of single cells from the biofilm.
Associated, multiple micro colonies emerge which are spatially distributed to optimize the hydrolysis of substrate to the end products- in the rumen,VFA and methane, and in the waste water treatment or biodigestors the end products are mainly methane and carbon dioxide. In summary, primary colonizing bacteria, that find a high concentration of their preferred substrate proliferate to form micro colonies and secrete high concentration of hydrolytic and other enzymes ( Cheng et al 1995; Sagiinala et al 1997) End product release seem to attract other organisms to the site and the EPS produced by the resident organisms facilitates and orders the attachment of these end product users( McAlister and Cheng 1996)
The close proximity of microbial species and enzymes appears to allow microbial cells to achieve a maximum net gain rate. Maximum net gain rate is defined by gross energy gain rate (derived from hydrolysate diffusion towards the cell) minus energy cost rate (represented by enzyme diffusion away from the cell; Vetter et al 1998). Microscopic studies by Akin (1976) indicated that the distance between cellulolytic rumen bacteria and the cellulose substrate surface is between 10–100 nm ensuring a high efficiency of cellulose hydrolysis.
Like enzymes immobilized in the ECP matrix, enzymes bound directly to the surface of feed particles are capable of cellulose hydrolysis (Song et al 2005). A biofilm growing on cellulose leaves pits in the surface layers (Cheng et al. 1981; Song et al 2005). These pits were produced by rod-shaped bacteria that have the same width as the grooves. The conformity of the pits on the feed particles to the bacterial shape indicates that cellulose is solubilised by cell surface- bound hydrolytic enzymes and not secreted enzymes held in the ECP-matrix. The physical attachment of the initial colonizing and hydrolyzing bacteria on the surface of fragments of plant particles is common and a typical phenomenon in rumen digestion of structural plant components (Cheng et al. 1981).
Biofilm with high levels of physiological efficiency was always found to be composed of complex multiple species (Stoodley et al 2002) as occurs in the rumen (see Edwards et al 2008; McAlister et al 2008; Firkins et al 2010). For example multi-species biofilm cultures appear to produce 70% more cellulose enzyme than single-species biofilms (Gutierrez-Correa and Tengerdy1999; Gutierrez-Correa and Villena 2003). In the case of solid substrate such as plant structural components, multi-species synergy, becomes highly important since sequential pathways or processes are involved and made efficient by the close association and spatial distribution of a number of cooperating microbial species within a layered biofilm matrix.
Direct microscopic examination of rumen contents has shown that cellulose degraders are the first organisms to colonize feed particles and then other species adhere to form a digestive consortium, resulting in biofilms with multi-species layers (McAllister et al. 1994). These microorganisms obtain their preferred or specific substrates that are produced locally as the end products the energy metabolism of close colony of a different species of micro organisms. The soluble components in the overall pathways of polymer breakdown thus move through the EPS matrix away from the surface of the plant particle (McAllister et al. 1994). The efficiency of this digestive consortium depends on how well the constituent microbes remove their preferred substrates because lowering the concentrations of products of the primary cellulose digestion relieves feed back inhibition of the initial reactions in the overall pathway. For example lowering of butyrate concentrations relieves inhibition of cellulose hydrolysis ( see Costerton 2007). Maintenance of a low hydrogen concentration within the biofilm matrix allows reduced cofactors to re- oxidized and drives the rate at which the cellulolytic process proceeds (Cheng et al. 1980).
In the absence of other high affinity electron acceptors such as nitrate, sulphur and some metals (e.g. Fe++), methanogenesis by reduction of carbon dioxide by hydrogen is critical for this sequential solid substrate degradation to proceed. Acetogenesis (Mackie and Bryant (1994) ( reduction of hydrogen to acetic acid presumably in the biofilm) in some situations also accomplishes the same objective of maintaining the hydrogen concentration at very low concentrations ( e. g. in the fermentative fore gut of marsupials (see Kempton et al 1976). Methanogenic archae/bacteria (and presumably acetogens) are generally found to strategically reside in the biofilm growing on solid substrate surface rather than dispersed in the supernatant, in spite of them being dependent on soluble substrate (Cheng et al. 1981). Colonies of methanogens are normally at the top of the biofilm structure with multiple species. This is usually on the side opposite the substrate (Song et al 2005) facilitating sequential degradation of polymers such as polysaccharides and proteins and thermodynamically favoring syntrophic breakdown of the organic matters (Liu et al 2002), with hydrolytic bacteria attached to the insoluble organic compound, followed by a fermentative bacteria layer responsible for the release and degradation of soluble products derived from the hydrolytic products, and a methanogenic colonies situated where they efficiently maintain hydrogen concentration low and release of VEA ( as in the rumen) or for complete mineralizing when the end products are methane and carbon dioxide as in biogas production (Schink 1997).
In the case of biogas production in digesters a high intermediate product concentration such as hydrogen , butyrate or acetate, can halt the whole anaerobic conversion reaction because the Gibbs free energy change for the anaerobic reaction does not make up for the minimum energy requirements for cell growth and maintenance (Jackson and McInerney 2002).
In water with low levels of particulate matter or where the organic matter is soluble such as in some sewage systems or even the liquid fed ruminant , biofilm formation is prevented from lack of inert surfaces and bioflocs may replace biofilms so as to enable a short physical distance between layered cooperating syntrophic species that drives the breakdown of polymers. One example occurs in macroscopic bioflocs that appear in sewage treatment plants where surfaces are of limited availability (Thiele et al. 1988). Bioflocs consist of diverse bacterial species embedded in a secreted ECP matrix. In these pseudo biofilm, the hydrogen producers and users are much closer to each other than if free floating in the bulk fluid, allowing efficient and rapid electron transfer. More than 90% of the hydrogen generated was consumed in the biofloc because of the of the multi-species synergy (Conrad et al. 1985). What appears now to be bioflocs were observed in rumen fluid of sheep fed a high molasses diet low in fiber (see Rowe et al 1979).
The preponderance of biofilm degradation of insoluble plant particulate matter undoubtedly allows these to be digested at a rate that allows ruminants to ingest sufficient feed materials to maintain and produce. The necessity of the biofilm mode of fermentative digestion indicates the important aspect of the hydrogen concentration and the role of the syntrophic microbes in rapidly capturing hydrogen as it is produced and maintain hydrogen below any level that has feed back inhibition of the re-oxidation of reduced cofactors in the parts of the overall pathways involved.
In the absence of other high affinity electron accepting compounds and the microbes that use these, methanogenesis is a critical requirement of the rumen mode of digestion. The use of chemicals in feed to inhibit methanogens then becomes unacceptable because it will decrease any syntrophic actions and decrease feed digestion in the ruminant .
Biofilms in waste water sewage treatment works are generally attached to the sewer wall or other inert services and are large and readily studied. They tend to go through considerable change with time from initiation of the biofilm through the growth of specially distributed colonies of microbes. They are general in flow through systems that apply a force to the surface of the biofilm matrix and when mature pieces break off new colonization processes commence. In the rumen biofilms are rapidly formed in their initial development being discernable with-in minutes when leaf material for instances is immersed in rumen fluid even in vitro (see Cheng et al 1980; McAllister et al. 1996; Edwards et al 2007). However the rumen has a rapid turnover of contents relative to other systems discussed above. Also it has an additional complexity not seen in many ecosystems of the water treatment facilities. The feed particles are small and constantly mixed within the fluid by contractions of the rumen-reticulum and the biofilm formation, maturation and erosion from the plant materials must be effected by short ( during feeding) or long term ( between meals) bouts of rumination in which the particles are subjected to considerable grinding by the teeth. Mature stable multi species biofilms are resistant to detachment (McAlister et al 1994; Harrison et al 2005) which is consistent in that it protects the entrapped microbes against a range of attacks , However the rapid rate of particle breakdown in the rumen relative to other systems suggest that potentially rapid renewal of biofilm colonies are needed to facilitate solubilization of plant particle components which maybe a feature of the rumination cycle of feed in ruminants. ruminants. In rumen fluid structural lignocelluloses undergo enzymatic hydrolysis by various bacteria ( and fungal) that sequentially attach to the feed particle (Cheng et al 1980) and in sacco studies with fresh forage have shown that mastication, besides increasing the areas of exposed substrate for hydrolysis is associated with colonization by different microbes. The mix of organisms on particles of perennial ryegrass seen after about 4 hours, were different to the primary colonizing bacteria (Kim et al 2006). The role of rumination and chewing in biofilm function and structure is explored more in the following discussion.
Ruminants use mouth and tongue to harvest
forages during grazing or to consume supplied feedstuffs. Cattle gather forage
during grazing by wrapping their tongues around the plants and then pulling to
tear the forage into the mouth from that anchored by the pasture roots and
following a short chewing period swallow the mouth full. On average, grazing
cattle take from 25,000 to more than 40,000 prehensile bites to harvest forage
each day (see Saras-Johansson undated). Depending on climatic and pasture
quality, in general they spend more than one third of their time grazing, about
one-third of their time ruminating, and the rest with no obvious activity.
However the patterns are different for different feed materials.
The roof of the ruminant mouth is a hard palate without incisors. The lower jaw
incisors work against this hard dental pad. The incisors are wide with a
shovel-shaped crown. Premolars and molars match in there opposing sites between
upper and lower jaws. These teeth are then able to crush and grind plant
material during initial chewing and particularly during the rumination cycle. Alkaline saliva aides in chewing and
swallowing and also has a buffering effect maintaining pH in the range from
6-7. During and following chewing, forage and feed
is mixed with a saliva to form a bolus which is propelled from the mouth by
contractions and pressure differences c down the esophagus to the reticulum
where it then enters the rumen.
Ruminants eat rapidly, usually swallowing much of their fibrous feed as large
fragments to be forced back from the rumen into the mouth sometime after the
feeding cycle is complete. In the process of rumination forage and other feeds,
having been immersed in rumen contents, are forced back to the mouth for further
chewing and mixing with saliva. This bolus is then re-swallowed and re enters
the reticulum. The solids in the bolus slowly move back into the rumen where
fermentation proceeds. In the formation of the bolus most of the liquid/small
particles are expressed and are positioned so that they precede the bolus when
this is swallowed, the liquid, small particles and detached microorganisms drain
into the omasum and then abomasum. The solid portion left behind in the rumen
typically remains within the cycle of rumination and floating in the rumen for
up to 48 hours where microbes can ferment the fibrous feedstuffs. This allows
separation of liquid and small feed particles, displaced microorganisms and
perhaps fragmented microbial biofilms to move to the lower parts of the
digestive tract.
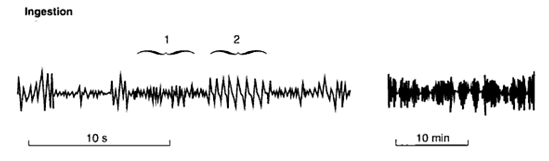
Chewing movements in feeding and rumination have different patterns and frequencies of jaw movements. Rumination has a more consistent frequency of both the jaw movements and intervals between bolus productions (Figure 2) than when the animal is initially eating. When the animal initially acquires feed, jaw movements are more irregular as shown in Figure 3. The irregular movements are due to minor interruptions and the alternation between prehension of feed and chewing sequences but most importantly even at this time a short rumination cycle may occur which could be a mechanism for seeding the newly acquired feed particles with microbes or more likely biofilm fragments.
 |
|
Figure 2. |
 |
| Figure 3. Jaw movement recordings transferred on
paper charts (from Baumont (2006) Recording at 15cm/h: Successive rumination cycles can be distinguished Recording at 30cm/min: Regularity of jaw movements in amplitude and frequency is observed |
Differences in rumination time are found between studies and between feed types. Welch and Hooper (1988) suggested that rumination on forage based diets occurs in shorter periods of 15-30 min at times during the feeding period and longer periods of two and six hours in the non feeding period. Ulyatt et al. (1986) and Ruckebush (1988) showed rumination of 5 and 9 hours but t shorter bouts between or during feeding were not included (see Welch and Hooper 1988; Baumont et al 2006).
The timing of rumination events, as discussed below, may be involved in the way (1) biofilm communities are separated from digesta and move into the gastric areas of digestion, (2) are disseminated onto newly acquired feed particles (3)disrupted and redistributed between particles or (5) rehabilitated on the surface of feed particles which maybe stripped of EPS containing sessile microbes, that may allow the biofilm to rejuvenate and /or change the spatial distribution of microbial species on return of the particle to the rumen in the bolus (see later).
Rumination includes periodic separation of liquid and small particles by squeezing the bolus with the tongue against the hard palate of the mouth. The suspended materials and liquid that is swallowed appears to be directed past the rumen and proceeds to the lower digestive tract potentially carrying biofilm fragments and microbial communities into the small intestines along with small particles with high levels of the least digestible components relative to the original forage.
Undoubtedly rumination has a profound effect on digestion of feed but this has not been specifically studied. Most researchers appear to subscribe to the concept that rumination is a process for increasing the surface area of solid materials to effectively provide greater access for fermentative organisms. However, it seems reasonable to suggest that rumination has profound effects on biofilm development, integrity and the separation of feed particles exhausted of the most of the digestible materials and is a mechanism for harvesting the microbes for digestion in the intestines.
The hypotheses presented here are that short bouts of rumination during feeding or shortly thereafter are mechanisms for transfer of rumen organisms between newly ingested feed and feed particles with mature biofilm. The longer periods of rumination between feed intakes are more associated with harvesting of the microbes within mature biofilm in the process of swallowing boluses.
From the above discussion, a set of conclusions can be developed. These are often implied from research with anaerobic ecological entities that are not the same as the rumen. These may be regarded as partially proven hypotheses or hypotheses waiting to be tested.
Biofilm related microbes are the primary organisms involved in the degradation of complex organic matter in the rumen
The biofilm consortia are distributed as sessile colonies spatially distributed to maximize the efficiency of energy utilization, This includes
Minimized distance between cooperating microbial colonies
Maintenance of minimal concentrations of end products that would initiate feed back inhibition of hydrolysis of, in particular, cellulose and hemi cellulose. This particularly applies to the concentration of H2
Low concentrations of hydrogen is maintained by biofilm colonies of methanogens that are critical in allowing oxidation of reduced co factors in the conversion of organic matter to VFA
In the absence of quantifies of high affinity electron acceptors other then carbon dioxide, methanogenesis is indispensable for efficient fermentative digestion in the rumen
From (4) there is no chance of mitigation of enteric methane production by inhibition of methanogens, without providing alternative electron accepting chemicals such as nitrate, sulphate and reduced iron oxides.
Even the addition of such compounds into the diet of ruminants must be linked to the growth, within the biofilm, of microbes that can use them in their energy metabolism.
Reduction of hydrogen by carbon dioxide as used by methanogens is energetically inefficient as compared to the reduction of nitrates and sulphate, removing any requirements for additional chemicals to directly inhibit methanogens.
The feeding and ruminating patterns of ruminants have major implications for the efficient formation and growth of the biofilm mode of digestion. Rumination is suggested to be a major mechanism for rapid colonization and re-colonization of feed particles with organized digesting and syntrophic colonizers that require to change as substrate availability changes with time in the rumen
Akin D E 1976 Ultrastructure of rumen bacterial attachment to forage cell walls. Appl Environ Microbiol 31:562–8
Anonymous 2011 Biofilm formation http://microbialecologynotes.wordpress.com/tag/microbial-ecology/
Baumont R, Doreau M, Ingrand S and Veissier I 2006. Feeding and mastication behaviour in ruminants. In: Feeding in domestic vertebrates: from structure to behaviour (ed: Bels, V.), 84-107. Cabi International. Wallingford, UK
Cheng K J and Costerton J W 1980 The formation of micrcolonies by rumen bacteria Can. J Microbiol 26, 1104-1113
Cheng K J, Fay J P, Howarth R E and Costerton J W 1980. Sequence of events in the digestion of fresh legume leaves by rumen bacteria. Applied and Environmental Microbiology 40, 613–625.
Cheng K J, Fay J P, Coleman R N, Milligan L P and Costerton J W 1981 Formation of bacterial microcolonies on feed particles in the rumen. Appl Environ Microbiol 41:298–305
Cheng K J, McAllister T A and Costerton J W 1995. Biofilm of the ruminant digestive tract. In Microbial biofilms (ed. H Lappin-Scott and JM Costerton),pp. 221–232. Cambridge University Press, Cambridge, UK
Chynoweth D P, Haley P, Owens J, Teixeira A, Townsend T, Xu Q and Choi H L 2003 Anaerobic composting for recovery of nutrients, compost, and energy from solid waste during space missions. In: Pullammanappallil P, McComb A, Diaz LF, Bidlingmaier W (eds) Proceedings of the 4th international conference of ORBIT association on biological processing of organics. ORBIT , Perth, pp 126–135
Conrad R, Phelps T J and Zeikus J G 1985 Gas metabolism evidence in support of the juxtaposition of hydrogen-producing and methanogenic bacteria in sewage sludge and lake sediments. Appl
Environ Microbiol 50:595–601
Confer D R and Logan B E 1997 Molecular weight distribution of hydrolysis products during biodegradation of model macromolecules in suspended and biofilm cultures.1. Bovine serum albumin. Water Res 31:2127–2136
Craig W M, Broderick G A and Ricker D B 1987. Quantitation of microorganismsassociated with the particulate phase of ruminal ingesta. Journal of Nutrition 117, 56–62
Costerton J. Wr. Cheng K J, Geesey G G,. Ladd T I, Nickel J G, Dasgupta M and. Marrie T J 1987. Bacterial biofilms in nature and disease. Annu. Rev. Microbiol. 41:435–464.
Costerton J. W. 1995a. Overview of microbial biofilms. J. Ind. Microbiol. 15:137–140..
.
Costerton J W,. Lewandowski Z, Caldwell D E, Korber D R and. Lappin-Scott H M 1995b. Microbial biofilms. Annu. Rev. Microbiol. 49:711–745.
Costerton J W 2007 The biofilm primer 1 Springer Series on biofilms Springer Berlin Heidelberg New York
Davey M E. and O’Toole G A. 2000 Microbial Biofilms: from Ecology to Molecular Genetics, Microbiol. Mol. Biol. Rev, vol. 64, pp. 847–867
Edwards J E, Huws S A., Kim E J and Kingston-Smith A. H. 2007 Characterization of the dynamics of initial bacterial colonisation of non-conserved forage in the bovine rumen FEMS Microbiology Ecology, 62 (3), 323-335 DOI - Online at: http://www.blackwell-synergy.com/doi/pdf/10.1111/j.1574-6941.2007.00392.x
Edwards J E, McEwan N R, Travis A J and Wallace R J 2004. 16s rDNA library-based analysis of ruminal bacterial diversity. Antonie van Leeuwenhoek 86, 263–281.
Edwards J E, Huws S A, Kim E J and Kingston-Smith A H 2007. Characterisation of the dynamics of initial bacterial colonisation of non-conserved forage in the rumen. FEMS Microbiology Ecology 62, 323–335.Microbiology 151, 717–725.
Edwards J E, Huws S A, Kim E J, Lee M R F, Kingston-Smith A H and Scollan N D 2008. Advances in microbial ecosystem concepts and their consequences for ruminant agriculture Animal, 2 (5), 653-660 DOI - Online at: http://dx.doi.org/doi:10.1017/S1751731108002164
Firkins J F 2010 Reconsidering rumen microbial consortia to enhance feed efficiency and reduce environmental impact of ruminant livestockproduction systems Revista Bras. Zootec 39 445-457
Flemming H C and Wingender J 2001 Relevance of microbial extracellular polymeric substances (EPSs)—part I: structural and ecological aspects. Water Sci Technol 43:1–8
Forsberg C W and Lam K 1977. Use of adenosine-5’-triphosphate as an indicator of the microbiota biomass in rumen contents. Appl. Environ. Microbiol. 33:528
Gutierrez-Correa M and Tengerdy R P 1997 Production of cellulase on sugar cane bagasse by fungal mixed culture solid substrate fermentation. Biotechnol Lett 19:665–667
Gutierrez-Correa M and Tengerdy RP 1999 Cellulolytic enzyme production by fungal mixed culture solid substrate fermentation. Agro Food Industry Hi-Tech 10:6–8
Gutierrez-Correa M and Villena G K 2003 Surface adhesion fermentation: a new fermentation category. Rev Peru Biol 10:113–124
Harrison J J, Turner R J, Marques L L R and Ceri H 2005 Biofilms: A new understanding of these microbial communities is driving a revolution that may transform the science of microbiology
American Scientist 93, pp. 508-515
Hungate R E 1966 The rumen and its microbes- Academic Press, Inc., New York.
Jackson B E and McInerney M J 2002 Anaerobic microbial metabolism can proceed close to thermodynamic limits. Nature 415:454–456
Kempton T J, Murray R.M and Leng R A 1976. Methane production and digestibility measurements in grey kangaroo and sheep. Australian Journal of Biological Science. 29, 209-214.
Kim E J, Edwards J E, Sanderson R, Kingston-Smith A H, Scollan N D and Theodorou M K 2006. Effects of mastication on temporal bacterial colonization and degradation of fresh perennial ryegrass in the rumen. Reproduction, Nutrition, Development 46, S19.
Krebs G 1987 The Rumen Ecosystem - Kinetics of Microbial Pools and Effects on Microbial Protein Yield' PhD dissertation. University of New England Armidale NSW 2605 Australia
Liu W T, Chan O C and Fang H H P 2002 Characterization of microbial community in granular sludge treating brewery wastewater. Water Res 36:1767–1775
MacLeod F A, Guiot S R and Costerton J W 1990 Layered Structure of Bacterial Aggregates Produced in an Upflow Anaerobic Sludge Bed and Filter Reactor Applied and Environmental Microbiology, June, p. 1598-1607.
Mackie R I. and Bryant M P 1994 Acetogenesis and the rumen: syntrophic relationships. In Acetogenesis. Ed: Drake, H.L. Chapman and Hall, New York. pp. 331-364.
McAllister T A, Bae H D, Jones G A and Cheng K J 1994. Microbial attachment and feed digestion in the rumen. Journal of Animal Science 72, 3004–3018
McAllister T A and Cheng K J 1996 Microbial strategies in the ruminal digestion of cereal grains. Anim Feed Sci Tech 62: 29–36.
Mayorga O L, Huws S A, Kim E J, Kingston-Smith A H, Newbold C J, Theodorou M K 2007.Microbial colonization and subsequent biofilm formation by ruminal microorganisms on fresh perennial ryegrass. Microbial Ecology in Health and Disease, 19 (1), p26 Conference on Gastrointestinal Function (CGIF), Chicago, USA, 16-18 April 2007
Minato H, Endo A, Ootomo Y and Uemura T 1966. Ecological treatise on the rumen fermentation. II. The amylolytic and cellulolytic activities of fractionated bacterial portions attached to the rumen solids. The Journal of General and Applied Microbiology 12, 53–69.
Nikolaev Yu A and Plakunov V K 2007 Biofilm-“City of Microbes “ or an analogue of multicellular organisms. Microbiology 76, 125-138
Nolan J V 2011 pers comm..
Preston T R and Leng R A 1985 Matching livestock systems to available resources in the tropics and subtropics. Penambul Books, Australia http://www.utafoundation.org/P&L/preston&leng.htm
Preston T R and Willis M B 1970 Intensive beef production. Pergamon Press, Oxford
Rappe M S and Giovannoni S J 2003. The uncultured microbial majority. Annual Review of Microbiology 57, 369–394
Rodríguez C J, González J, Alvir M R, Redondo R and. Cajarville C 2003 Effects of feed intake on composition of sheep rumen contents and their microbial population size British Journal of Nutrition 89, 97-103
Ruckebush Y 1988 Motility of the gastro-intestinal tract. In: The ruminant animal: digestive physiology and nutrition (ed. Church, D. C.) 64-108. Prentice Hall. Englewood Cliffs, USA.
Rowe J B, Loughnan M L ,Nolan J V and Leng R A 1979 Secondary fermentation in the rumen of a sheep given a diet based on molasses; British J Nutrition 41 393-397
Saras-Johansson M Undated Chewing behaviour of growing cattle for discussion http://stud.epsilon.slu.se/2823/1/saras_johansson_m_110616.pdf
Schink B 1997 Energetics of syntrophic cooperation in methanogenic degradation. Microbiol Mol Biol Rev 61:262–280
Song H, Clarke W P and Blackall L L 2005 Concurrent microscopic observations and activity measurements of cellulose hydrolyzing and methanogenic populations during the batch anaerobic
digestion of crystalline cellulose. Biotechnol Bioeng 91:369–378
Stoodley P, Sauer K, Davies D G and Costerton J W 2002 Biofilms as complex differentiated communities. Annu Rev Microbiol 56: 187–209
Sutherland I W 2001 Biofilm Exopolysaccharides: a Strong and Sticky Framework, Microbiology (UK), 2001, vol. 147, pp. 3–9.
Tengerdy R P and Szakacs G 2003 Bioconversion of lignocellulose in solid substrate fermentation. Biochem Eng J 13:169–179
Thiele J H, Chartrain M and Zeikus J G 1988 Control of interspecies electron flow during anaerobic-digestion—role of floc formation in syntrophic methanogenesis. Appl Environ Microbiol 54:10–19
Ulyatt M J, Dellow D W, John A, Reid C S W and Waghorn G C 1986. Contribution of chewing during eating and rumination to the clearance of digesta from the ruminoreticulum. In: Control of digestion and metabolism in ruminants (eds: Milligan, L. P, Grovum, W. L. & Dobson, A.) 498-515. Prentice-Hall, Englewood Cliffs, USA
Vetter Y A, Deming J W, Jumars P A and Krieger-Brockett B B 1998 A predictive model of bacterial foraging by means of freely released extracellular enzymes. Microb Ecol 36:75–92
Weimer P J, Russell J B and Muck.R E 2009 Lessons from the cow: What the ruminant animal can teach us about consolidated bioprocessing of cellulosic biomass Bioresource Technology 100 5323–5331
Wetzel R G 1991 Extracellular enzymatic interactions: storage, redistribution and interspecific communication. In: Chrorst RJ (ed) Microbial enzymes in aquatic environments. Springer, New York, pp 6–28
Wang and Chen 2009 Potential o-based biofuels production Appl. Microbiol. Biotechnology 83, 1-18
Welch J G and Hooper A P 1988. Ingestion of feed and water. In: The ruminant animal: digestive physiology and nutrition (ed: Church, D.C.) 108-116. Prentice Hall, Englewood Cliffs, USA
Wolin M J, Miller T L and Stewart C S 1997. Microbe–microbe interactions. In The rumen microbial ecosystem (ed. PH Hobson and CS Stewart), pp. 467–491.Chapman & Hall, London, UK