Pre- and post-natal exposure to duckweed affects its post-weaning familiarity and intake in Large White and Mong Cai pigs
Abstract
In Expt 1, offspring of sows (Large White and
Mong Cai breeds) that were
either never exposed to duckweed, or were exposed daily to duckweed throughout
pregnancy were tested to determine whether their experiences in utero
affected their rate of acceptance and intake of duckweed in later life. After
weaning, when the offspring were offered duckweed for the first time, the Large
White offspring more readily accepted and ingested duckweed if their mothers
had been exposed to duckweed during pregnancy. Mong Cai offspring also appeared to have learned about
duckweed in utero
but they ingested duckweed more
readily than Large White offspring, irrespective of whether or not their
mothers had been exposed to duckweed.
In Expt 2, sows of
both breeds and their offspring were given duckweed to eat each day when the
offspring were 3-5 weeks of age while other groups of sows and their offspring
were not exposed to duckweed. Offspring with and without pre-weaning exposure
to duckweed were then tested after weaning (as in Expt 1) to determine their
familiarity with and ingestion of duckweed. Both breeds accepted duckweed more
readily if they had been exposed to duckweed with their mothers in the
pre-weaning phase, but Large White offspring were slower to accept and ingest
duckweed than Mong Cai offspring.
The time taken for weaned pigs to accept and begin to ingest a newly offered food appears to depend on whether the food is familiar. Such familiarity may result from personal experience (including in utero experience) or learning assisted by watching or participating with social models such as mother when they ingest that food. We conclude that reductions in growth rate at weaning time when piglets are first offered solid foods, or subsequently due to frequent changes in diet, can be reduced if novel foods are given a salient flavour of a food eaten prenatally by mother or post-natally by mother and piglets together.
Introduction
In rural areas in
Duckweed (Lemma spp) is a high yielding aquatic
plant capable of both removing environmentally unfriendly minerals from waste
water (Leng et al., 1995; Leng, 1999) and supplying protein, minerals and other nutrients
for monogastric animals and also fish (Preston, 2000). However, when duckweed is offered for the first time
to pigs, those pigs (especially exotic
breeds) exhibit neophobia and do not ingest it for up
to 7 days (T.R. Preston, personal communication). When animals are seen to be
reticent to ingest novel foods (Domjan and Gregg, 1977) farmers may be discouraged from using such foods for
livestock production. Food neophobia is one of the
main constraints to the use of unconventional foods by exotic monogastrics and ruminants, especially the goat in
Currently, Vietnamese government policy makers are promoting the idea that local agro-byproducts should be used as supplementary foods for livestock. There is a need to understand better the role of early-life training and/or addition of familiar flavours to unconventional foods in order to attract animals to the feeders and encourage them to begin to eat these foods with minimum delay.
Village farmers are generally unaware that neophobia and food avoidance by animals often occurs simply because the foods are unfamiliar and often conclude that foods are unsuitable for their animals or they feel they do not have the time or the patience to provide daily training sessions, possibly over several weeks, to train their animals to accept and ingest unconventional foods.
Bilko et al. (1994) have proposed that young
rabbits may learn about their mother’s diet from their experiences in utero, by
proximity to mother during suckling, or from their chemo-sensory responses to
their mother’s milk or to her faecal pellets
deposited in the nest. Galef and
Henderson. (1972) had earlier argued that an offspring can learn about
a food that its mother has ingested from food odours
and tastes that are transmitted via her milk. Young animals may also learn
about food the mother has eaten if it contributes to their mother’s body odour (
Young animals including sheep (Lobato et al., 1980; Nolan et al., 1995; Provenza, 1996) pigs (Tien, 2000), cats (Wyrwicka, 1993) and humans (Birch, 1998) also learn about foods in the pre-weaning phase by social interactions. A mother is a particularly effective social model for her offspring because she is closely related genetically and hence should respond more similarly to smell, taste and post-ingestive feedback than unrelated individuals (Chapple and Lynch, 1986).
We hypothesized that when a pregnant sow was given a supplementary food, the odour or flavour of that food might be sensed in some way by her fetuses enabling them to become familiar with its characteristics. If an offspring can learn in utero about foods sows eat during pregnancy, or during the pre-weaning phase, their resulting familiarity with the food cues could reduce their neophobia for solid foods during and after weaning. This could contribute to reduce the set-back in production often suffered by young pigs at weaning time as they are becoming accustomed to solid foods and, in later life, increase their intake in the short term after sudden introductions of unconventional foods.
This study was conducted with
young pigs to determine if their experience of a sow’s diet in utero (Expt 1), or their
post-natal exposure to food cues via milk or body odour,
or participation with the sow at meal-time (Expt 2)
would enhance their post-weaning acceptance and intake of duckweed. The study
was made with two breeds of pig, i.e. native (Mong Cai) and exotic (Large White) breeds in
Materials and
methods
General
The study was carried out at the
state Pig Breeding Research Institute near to
The pigs were housed in concrete
sheds with concrete floors and open sides and they were raised according to the
normal management procedures in use at the Institute. All pigs were given a commercial diet
(cassava, rice bran, dried fish-meal and minerals/vitamins; 3098 kcal or 12.8
MJ metabolisable energy/kg; 140 g crude protein/kg)
supplemented with freshly harvested sweet potato forage as is the traditional
practice in the region. When required, duckweed was harvested from ponds within
25 m of the furrowing shed. In the post-natal, pre-weaning phase, sows and
piglets were offered the same commercial diet (2.0 kg/d) supplemented with
freshly harvested sweet potato leaves (700 g/d). Piglets were weaned at about
10 weeks’ of age when the Large White piglets were about 19 kg and Mong Cai about 12 kg. After weaning the
young pigs were given the same commercial diet up the time of testing at about
3 months of age.
Experimental
design
In this study, 32 young piglets
(16 Large White and 16 Mong Cai) were obtained from
the litters of sows that had ingested duckweed throughout pregnancy (Expt 1) or piglets that had experienced their mother ingesting
duckweed in the post-natal, pre-weaning phase (Expt
2). Duckweed was used as an unconventional food because it is a potential
source of good-quality protein for pigs in central Vietnam and did not form
part of the normal diet used at the Breeding Institute.
Experiment 1
Expt 1 was undertaken to test if information about
duckweed ingested by a sow during her pregnancy could be passed to her fetuses,
thereby influencing their post-weaning acceptance and intake of duckweed. Young
pigs were selected at random from litters of 2 sows from 2 breeds (Mong Cai and Large White) to make four
groups each of 4 pigs. The first two treatment groups were selected from the
litters of two Mong Cai sows
with litter sizes of 12 and 10 piglets, respectively. Groups 3 and 4 were
selected from Large White sows with litters of 14 and 9 piglets, respectively.
Group 1 sows and their fetuses were exposed to duckweed (i.e. ‘exposed in utero’) during
their pregnancy until three days before parturition, but not subsequently,
whereas Group 2 sows were given the same management, but were not offered
duckweed at any time (‘non-exposed’).
Throughout their pregnancy, sows
in Group 1 were offered fresh duckweed (initially at 200 increasing to 500
g/head per day in the last days’ of offering) between 06.00 and 07.00 h after
being fasted overnight, followed by 1.5 kg/d of the commercial diet described
above. All 4 sows gave birth in
European steel farrowing pens in the same shed
between 3rd and 7th March 2000. The piglets were left in
the farrowing pen with their mother for about 5 h
after birth and then the sow and her litter were moved to a single pen in a
nearby shed.
In this experiment, efforts were made to reduce the possibility that information about the characteristics of duckweed ingested by the sows during pregnancy might be passed to the piglets in the early post-natal period, e.g. by the presence of residual duckweed odor in the area where the pigs were born. To eliminate this possibility, duckweed was removed from the diet of the sows on day 111 of pregnancy (started by artificial insemination) which was 3 days before they were due to give birth. The pens, feeders and surroundings were cleaned twice daily with water delivered under pressure via a hose, including in the 3-day period before piglets were born.
At three months’ of age and
about 3 weeks after weaning, the offspring in Group 1 and Group 2 were exposed,
for the first time post-natally, to duckweed in a
daily testing process aimed at determining their familiarity with duckweed.
Groups 1 and 2 (Mong Cai
breed) were managed and tested using the same procedures as Groups 3 and 4
(Large White breed).
Experiment 2
Expt 2
was undertaken to evaluate if pre-weaned piglets that had been exposed to
duckweed from 3 to 5 weeks’ of age (by participating when their mothers were
ingesting duckweed, and/or by sensing duckweed characteristics in their
mothers’ milk) would exhibit an increased preference for, and a higher intake
of duckweed than those without any experience of duckweed when tested at 3
months’ of age. Groups A and B were selected at random from litters of 2 Mong Cai sows (litter sizes of 12 and 13, respectively) and
Groups C and D were selected from the litters of 2 Large White sows (litter
sizes of 12 and 10 piglets, respectively).
For Group A, 3- to 5-week-old piglets could observe and participate while their mothers ingested duckweed (i.e. ‘exposed’) and could potentially learn about duckweed directly, or via duckweed cues in their mothers’ milk. The training procedure was as follows. After an overnight fast in the holding pen, each mother was moved with her piglets to a pen where all the animals were offered fresh duckweed for 15 min (at 800 gradually increasing to 1000 g/litter per day in the last week of training). The piglets had the opportunity to observe and participate while their mothers were eating. After training, the animals were again offered their normal commercial diet supplemented with freshly harvested sweet potato forage for the rest of the day. Three months later, after weaning and in the absence of their mothers, the piglets were offered fresh duckweed in a test that was similar to that used in Expt 1. In contrast, Mong Cai pigs in Group B had no exposure to duckweed at any time before the commencement of the post-weaning testing (‘non-exposed’).
Groups C and D (Large White
breed) were managed and tested using the same procedures as Groups A and B
(Mong Cai breed).
Testing area
The testing area had separate test stations (5 m x 1 m) that provided enough space for up to 10 young pigs to be present at 10 separate feeders and to eat test foods side by side. Woven bamboo was used to fabricate the sides and compartment dividers that allowed the pigs to see each other during testing. Two fenced pathways were made to guide the animals in Expts 1 and 2 from covered houses (where pigs were housed overnight) to the testing area.
Testing
procedures
After weaning at about 10 weeks’
of age, young pigs were tested at 12 weeks’ of age to determine their
acceptance and voluntary intake of duckweed in a daily 15-min opportunity to
ingest duckweed. All animals were adapted to the testing facilities and
procedures for 5 days before the testing commenced. Sham tests were undertaken
using freshly harvested sweet potato leaves. In Expt
1, after an overnight fast, Groups 1-4 were allocated to the testing station at
05:30 to 06:00; 06:15 to 06:45; 07:00 to 07:30 and 07:45 to 08:15 h,
respectively. Each pig was offered 300‑600 g duckweed, depending on its
individual history of ingesting the test food in the 15-min test. Some young pigs
did not ingest any duckweed for some days and, during the pre-ingestion phase,
the incidences of ‘nosing’ (bring the muzzle close to the food) and ‘muzzling’
(making contact between the muzzle and the test food) were recorded. The
testing continued until the daily intakes for each group were approaching a
plateau value. The intake of duckweed by each pig was determined by weighing
the duckweed in each feeder before and after each 15-min test. All feeders were
thoroughly cleaned after each test.
For Expt 2,
the schedule for testing pigs in Groups A-D was the same as for Experiment 1.
Statistical analysis
Analysis of variance was
performed using the general linear model (GLM) procedure of Mintab
version 12 (1998). Comparisons with a confidence level of 95 % were used to
determine the effects of behavioral treatments between groups.
Results
Experiment 1 (training in utero)
In early pregnancy, the sows adapted within one week to eating fresh duckweed. Thereafter, the intake of commercial food by the Mong Cai sows (Group 1) was 1.5 kg/d and intake of duckweed was 0.7 kg/d. Corresponding values for the Large White sows (Group 3) were 2.0 kg/d and 0.85 kg/d. At the start of post-weaning testing, the young Mong Cai pigs weighed about 15 kg and their weight increased to 18 kg during the 30-day period of testing (weighing accuracy ±1-2 kg). Corresponding live weights for the Large White pigs were 23 and 27 kg. The intakes of duckweed by the pigs during the post-weaning are given in Figs 1A and 1B.
Once they began to ingest duckweed, intakes of pigs increased progressively towards a plateau intake, starting at about 25 days for the Large White and about 20 days for the Mong Cai pigs. By the time both the trained and untrained groups of Large White pigs had reached a plateau value at 25 days, the trained group had eaten a total of 5.99 kg duckweed during the testing period, an amount that was significantly (P<0.05) more than the untrained group that had ingested a total of 4.47 kg. For the Mong Cai pigs, the cumulative total intakes of trained and untrained pigs after 20 or 25 days of testing did not differ significantly (means 3.61; 5.74 kg, respectively). The mean plateau intake in the daily tests of the Large White pigs and for the Mong Cai pigs was similar (about 45 g/kgW0.75 see Figs 1A and 1B).
Some of the ‘non-exposed’ pigs did not ingest
duckweed until Day 9 of testing. In the 15-min tests, young Mong Cai pigs, in general, started to eat sooner (within 3-5 d)
than young Large White pigs (within 5-8 d).
Unexposed Mong Cai pigs did not start eating
duckweed until Day 5 of testing whereas exposed piglets started eating on Day
3. Large White pigs in the non-exposed groups ate nothing until Day 8 whereas
pigs in the exposed group started to eat on Day 5.
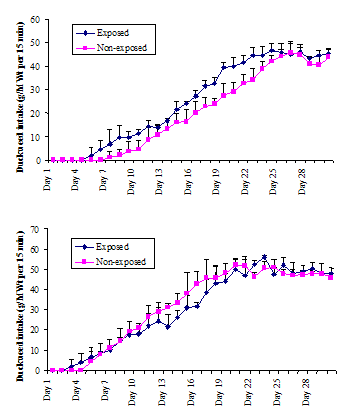
Figs 1A and
1B. Intake (g/animal) of duckweed during daily
post-weaning tests by (A) young Large White pigs and (B) young Mong Cai pigs raised by sows that had either ingested duckweed
throughout pregnancy (i.e. ‘exposed’ piglets had had the opportunity to learn
in utero) or ‘non-exposed’ piglets raised by sows
that had never ingested duckweed. Results are shown as means ± SD for pigs per treatment
Experiment 2 (training via observation)
Intakes of duckweed by pigs trained during the pre-weaning period, either by observing and possibly participating when their mothers were ingesting duckweed (direct training) or via mothers’ milk (indirect training) differed significantly from the intakes of pigs with no previous exposure to duckweed (see Figs 3A and 3B). The Mong Cai piglets with previous exposure to duckweed, investigated duckweed and ate small amounts of it on the first day of testing whereas the similarly exposed Large White pigs did not eat until Day 3 of the test. The unexposed Mong Cai pigs began eating on Day 4 whereas the unexposed Large White pigs did not begin to eat duckweed until Day 10 and did not reach a plateau intake by Day 21 when testing ceased. After commencing to eat the duckweed, all animals, i.e. those with previous experience and both control groups without previous experience, ate significant quantities of the duckweed, and their daily intakes increased progressively towards plateau values that were similar to those of pigs in Expt 1.
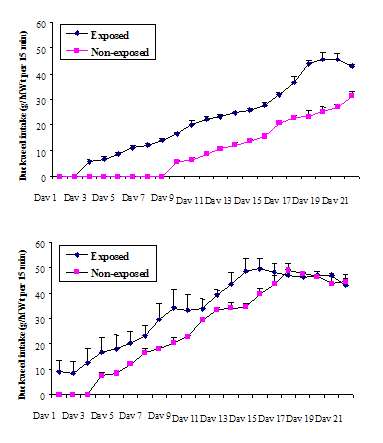
Figs 3A and 3B. Intake of duckweed (g/head per 15-min test)
by young pigs offered duckweed at 3 months of age after having been exposed
with mother to duckweed in the post-natal, pre-weaning period (‘exposed’) or
never exposed to duckweed (‘non-exposed’ ) (A - Large White pigs; and B - Mong Cai pigs).
Again Mong Cai pigs reached their plateau intake earlier (after about 18 d) than Large White pigs (> 21 d). In the 21 days of testing, the cumulative intake of exposed Large White pigs was 5.33 kg which was significantly (P<0.001) more than the cumulative intake of their unexposed counterparts (2.59 kg). The plateau values for intake during testing were reached earlier in the Mong Cai pigs (at about Day 14 for the exposed pigs and Day 17 for the unexposed pigs): by Day 17 the cumulative intakes of the exposed pigs (5.82 kg) was significantly (P<0.001) higher than for the unexposed pigs (4.63 kg).
Discussion
Neophobia leading to food avoidance is probably an innate survival mechanism. A low intake in the period immediately after animals are offered an unfamiliar food, allows them to test the food and experience any potentially life-threatening post-ingestive effects. They can then minimise their food intake to limit their absorption of anti-nutritional factors and/or other harmful substances or, if the food is safe and nutritious, increase their intake and obtain the nutritional benefits (Wong and McBride, 1993). This is a form of learning by ‘trial and error, i.e. the animal learns from its own experiences (Provenza and Balph, 1987). Familiarity with the sensory properties of foods helps overcome neophobia so that new foods that are familiar will be eaten more quickly and in larger amounts than foods that are novel. However, extinguishment of neophobia takes time during which food intake and production suffer. Moreover, food neophobia and reduced intake still occurs even when a new food is nutritious and non-toxic.
The indigenous Mong Cai pigs appeared to exhibit less neophobia and therefore the effects of exposure to duckweed both in utero and during the pre-weaning period were less obvious than for the exotic Large White breed. The intensity of food neophobia may be heritable and may vary between pigs of different breeds. Mong Cai pigs may have evolved with a less intense neophobic response to novel foods because of their nutritionally harsh environment and the balance of risk of death by poisoning against the risk of death from starvation. Large White pigs, on the other hand, that have become adapted to commercial diets may be more neophobic when confronted with unusual foods even though their risk of being poisoned by commercial foods is probably lower that for the more feral Mong Cai breed. Growth and production of Mong Cai pigs will therefore also be less affected by frequent introductions of new foods into the diet than would Large White pigs but both breeds can be expected to suffer a check in production when unfamiliar foods are offered.
Expt 1 has provided the first evidence, as far as we are aware, that the fetal pig can learn about foods its mother has eaten during her pregnancy and, without any post-natal experience of that food, exhibit familiarity with it in later life. It is not clear how such learning could occur. It seems probable that the fetus would have to sense a flavor or odor characteristic of duckweed, or a metabolite of duckweed, that was not produced by the remaining components of the diet given to the sows whose litters provided pigs for the control group. In rats, chemical cues (garlic) in mother’s blood can cross the placenta or diffuse into the amniotic fluid and be sensed by her offspring (Hepper, 1989). Flavours of foods a mother eats are transferred to amniotic fluid and the fetus, which drinks the fluid, and has functional olfactory bulbs in late pregnancy (Mennella et al., 1995) and Schaal et al. (1998) showed that human babies exhibit a highly selective neonatal recognition of their own amniotic fluid odor, and this affects their later behavior. Mennella. (2001) concluded that prenatal experiences of human fetuses when their mothers were ingesting carrots, perhaps mediated through flavour of amniotic fluid, lead to their greater post-weaning acceptance of carrot-flavoured foods relative to foods without carrot flavour. Thus studies of human and other animals lend support to our results reported here for the pig which indicate that chemo-sensory information about duckweed detected in utero can affect its post-weaning familiarity and subsequent ingestion.
Because of the novelty of the findings in Expt 1, we looked closely at possible ways by which the young piglets might have learnt about duckweed in the early post-natal period rather than in utero, even though duckweed was removed from the diets 3 days before farrowing. Could these piglets have gained information post-natally about the mother’s pre-natal diet by, for example, residual smells in the shed, or residues of food or excreta in the pens where their mothers had been housed? We consider it unlikely that residual duckweed cues could have persisted in the post-natal period, in light of the twice daily washing of the pens and feeders in the 3-day pre-farrowing period, and in any event, any residual cues would probably have also affected the piglets in the control groups that were born in the same shed.
It is possible that piglets might have been exposed post-natally to characteristics of duckweed via mother’s body odour, although this seems relatively unlikely given the 3-day pre-natal period when mothers were not ingesting duckweed. Future studies are planned in which newly born piglets will be cross-fostered to investigate this possibility. Another possibility is that cues from duckweed (chemical substances in duckweed, or metabolites of such chemical substances, with a characteristic taste or odour) were stored in the sow’s fat depots during pregnancy and released into her milk in the early part of lactation when she was drawing on energy in her body reserves. These duckweed cues might then have been detected in the milk ingested by the young pigs, making these cues familiar when they were experienced again during the post-weaning tests. This suggestion is compatible with the positive responses of the piglets to their post-natal learning about duckweed demonstrated in Expt 2.
In Expt 2, we examined the hypothesis that young pigs can learn about duckweed by detecting chemo-sensory duckweed cues in milk produced by their mothers while ingesting duckweed, or by social transmission of information about duckweed gained from their mother on the occasions she ingested duckweed. There is considerable evidence from species other than pigs that both types of information transfer can affect subsequent food familiarity in weaned animals of a number of species including the pig. For example, studies of rats (Galef and Henderson, 1972) and sheep (Nolte and Provenza, 1992) have shown that flavour cues in mother's milk can determine the feeding preferences of offspring after weaning.
Galef (1974) also concluded that offspring learn from their mother and from other animals by social interaction. A study by Madsen (1977) with pigs is of interest in this connection. He studied food preferences in 2100 young pigs, hypothesising that they might learn about flavours of foods being eaten by the sow either when they suckled or by being exposed to the food when the sow was ingesting it. He concluded that there was transmission of food preference, but only when the salient flavour was present in the diet of both the sow and the offspring.
Madsen (1977) also commented: ‘Weight loss at weaning, usually ascribed to stress, could be reduced if the flavour of the food was the same as that of the food eaten previously by mother and piglets’. This statement gains support from the results of the present study and from work by Bonomi et al. (1991). The latter workers found that offspring of sows given mixed foods that included a commercial flavour compound (containing essential plant oils with 0.04% vanillin), and also given access to food containing the same flavour from an early age, gained more weight after weaning at 2-3 weeks of age, and converted food to gain more efficiently than pigs raised under similar conditions without exposure to the flavor.
Conclusion
We conclude that young pigs can learn about a food in utero or by a combination of the effects of suckling and social interactions when their mother ingests that food in the post-natal, pre-weaning period, and so be more familiar with that food when they first encounter it in the post-weaning period. In general, Large White pigs exhibit stronger neophobia and require more training than Mong Cai pigs to overcome it. Animals that recognize characteristics of the food from previous experience of it before birth or in the post-natal, pre-weaning period, exhibit more investigatory behaviour and commence eating it sooner than those without experience. When unconventional foods are offered to pigs or when the diet is changed frequently, the short-term depression in intake and production that often results can be reduced if the animals have been managed to have previous pre-natal or post natal experiences of those foods.
Acknowledgements
This work was supported by the International Foundation for Science (IFS).
References
Bilko, A., V. Altbacker and R.
Hudson. 1994. Transmission of food preference in the rabbit- the means of
information transfer. Physiol. Behav. 56: 907-912.
Birch, L. L. 1998. Psychological influences on the childhood diet. J. Nutr. 128 Suppl: 407S-410S.
Bonomi, A., A. Sabbioni and Pantiglioni. 1991. Remarks and observations on the use of flavored diets in the early weaning of piglets. Annali della Facolta di Medicina Veterinaria, Universita di Parma 10: 249-264.
Chapple, R. S. and Lynch, J. J. 1986. Behavioral factors modifying acceptance of supplementary foods by sheep. Res. Develop. Agric. 3: 113-120.
Domjan, M. and Gregg, B. 1977. Long-delay backward taste-aversion conditioning with lithium. Physiol. Behav. 18: 59-62.
Galef. B. G. Jr. and Henderson. 1972. Mother’s milk: A determinant of the feeding preferences of weaning rat pups. J. Comp. Physiol. Psychol. 78: 213-219.
Galef. B. G., Jr. 1974. The social transmission of acquired behavior. In Mother-Young relationships: The Transfer of Information: pp. 155-160. Boston, Massachusetts: Plenum Publishing Corporation, New York.
Hepper, P. G. 1989. Foetal learning: Implications for psychiatry? Br J Psychiatry 155: 289-293.
Leng. R. A. 1999. Duckweed. Rome: FAO.
Leng. R. A., and Stambolie, J. H. 1995. Duckweed-potential high protein feed resources for domestic animal and fish. Livest. Res. Rural Develop. 7, no. 1.
Lobato, J. F.P., Pearce, G. R. and Beilharz, R. G. 1980. Effect of early familiarization with dietary supplements on the subsequent ingestion of molasses-urea blocks by sheep. Applied Animal Ethology 6: 149-161.
Madsen, F. C. 1977. Development of feed preference in young swine. Feedstuffs. USA. 49: 25.
Mennella, J. A. 2001. Regulation of milk intake after exposure to alcohol in mother’ milk. Alcohol Clin. Exp. Res. 25.
Mennella, J. A., Johnson. A, and Beauchamp. 1995. Garlic ingestion by pregnant women alters the odor of amniotic fluid. Chem. Senses 20: 207-209.
Nolan, J. V., Hinch, G. N. and Lynch, J.
J. (1995). Feeding behavior and nutrient intake in
ruminants. Recent Advances in
Nolte, D. L. and Provenza. F. D. 1992. Food preferences in lambs after exposure
to flavors in milk. Appl. Anim.
Behav. Sci. 32: 381-389.
Nguyen Van Lai. 1988. On-farm comparison of Mong Cai and Large White pigs fed
ensiled cassava root, rice bran and duckweed. Livest. Res. Rural Develop.
10, no. 3:1-6.
Nguyen Van Lai and Rodriguez. 1988. Digestion and metablism
in Mong Cai and Large With
pigs having free access to sugar cane juice or ensiled cassava root
supplemented with duckweed or ensiled cassava leaves. Livest. Res. Rural Develop. 10:
1-7.
Preston. T.R. 2000, Livestock Production from Local
Resources in an Integrated Farming System: A Sustainable Alternative for the
Benefit of Small Scale Farmers and Environment. In
‘Sustainable Livestock Production on Local Feed Resources’. Ed. Preston,
T. R. In: Sarec
Proceeding, Saigon, Vietnam January 2000. Pp: 15-21. Agricultural
Publishing House. Vietnam.
Provenza. F. D. 1996. Role of learning in food preferences
of ruminants: Greehalgh and Reid revisited. In ‘ Ruminant Physiology: Digestion, Metabolism, Growth and
Reproduction. Proceedings VIII International Symposiumon
Ruminant Physiology’ (W. von. Engelhardt, S. Leonhard-Marek, G. Breves, et al., eds), pp. 233-247. Ferdinand Enke Verlag, Stuttgart. Germany.
Provenza, F.D., and Balph, D.F.
1987. Diet learning by domestic ruminants: Theory, evidence and practical
implications. Applied Animal Behavior
Science (18) 211-232.
Schaal, B., Marlier,
L, and Soussignan. R. 1998. Olfactory function in the human fetus:
Evidence from selective neonatal responsiveness to the ordor
of amniotic fluid. Behav. Neurosci. 112: 1438-1449.
Tien. D. V. 2000. Current practices, potentials and
constraints of livestock production and solutions to develop the sustainable
livestock production in the Phongdien district, Thua Thien Hue province and to
increase income of farmers. In Livestock Management in the Threatened Flood Areas
of Thua Thien Hue. Annual Report of Rural Development Programme
October 2000. Pp. 2-6 TTH Press. Vietnam.
Wong, R., McBride, C.B., 1993. Flavor neophobia in gerbils (Meriones unguiculatus) and hamsters (Mesocricetus auratuss) and hamsters (Mesocricetus auratus). Quart. J. Exp. Psych., Sect. B. 46, 129-143.
Wyrwicka, W. 1993. Social effects on development of food preferences. Acta Neurobiol. Exp. (Warsz) 53: 485-493.