|
|
|
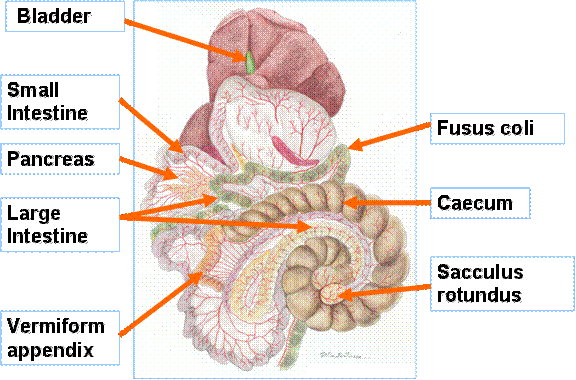
Figure 1: The digestive tract of the rabbit |
|
MEKARN Workshop 2008: Organic rabbit production from forages |
Rabbits are hind gut fermentors that, because of their high surface to body weight ratio, have a relatively high maintenance energy requirement. Rabbits given concentrate based diets (energy dense) need to consume 5% or more but with forage alone this is increased to over 8% of body weight on a dry matter basis.
Rabbits efficiently utilise fibrous feed by courtesy of their feeding and digestive strategies. They are highly selective when given forage, free choice and in quantity. Their digestive strategies include initial enzymic digestion in acidic followed by alkaline medium of the stomach and small intestines respectively, followed by fermentation of feed residues in the caecum large intestines.
The rabbit’s caecum is the largest organ in the digestive tract and it develops over about 1 month from birth to be truly functional. Initially young rabbits depend for their nutrition on milk which is high in fat and protein but contains only small amounts of lactose. The C8 and C10 fatty acids in milk fat are bacteriostatic and protect the young animal from invasion by detrimental organisms. As milk intake diminishes towards weaning the organisms that develop in the gastrointestinal tract particularly in the caecum depend on the type of carbohydrate and the protein flow into the caecum. Recent research has demonstrated an adverse effect of high protein diets on survival of rabbits after weaning and throughout the fattening period.
The rabbit has the ability, through specialised musculature of the large intestine, to direct slowly fermented fibrous feed into the colon for excretion in the hard faeces. The same mechanism separates small particles (more digestible fiber) and soluble components of digesta leaving the ileum into the caecum where it remains in a buffered medium supporting bacterial growth. The rabbit quickly “sieves” indigestible or slowly fermentable fibers which would slow its feed intake were they to enter the caecum. Hard faeces are produced some 4 hours after a meal.
The caecum is relatively large and the products of fermentation are similar to the products produced in the rumen of cattle and sheep- that is volatile fatty acids (VFA) and microbial cells. A number of reasons are put forward for the microbial growth being highly efficient in the caecum of rabbits on forage-based diets yielding a higher ratio of cells to VFA than may occur in fermentation in the forestomach of ruminants. Considerable VFA are apparently absorbed by the rabbit but the availability of the essential amino acids of microbial protein depends on the subsequent consumption of soft faeces or caecotropes.
Caecotropes are formed from the contents of the caecum as they pass through the large intestine and colon of the rabbit. The feed residues and culture medium are voided from the caecum about 8hours after a meal. The digesta with its microbes are formed into pellets by dehydration and coated with a mucous membrane as they pass along the colon. These soft pellets or caecotropes are retrieved as they pass from the anus and are swallowed by the animal without disrupting their membranes; they are also buffered towards a neutral pH . These pellets enter the acidic stomach and reside in the fundus portion ( which is close to neutral pH) for several hours. The membrane and the buffered contents maintain the pellet contents at close to neutrality. They finally disintegrate with increasing feed intake and the contents are then subject to gastric and intestinal digestion.
The concept is floated here to explain why the process of caectrophy may be a highly efficient strategy to utilise microbial cells produced in the hind gut. Recent studies have indicated the involvement of a group of enzymes termed lysozymes in the degradation of bacterial cells. Many secretions of the body contain lysozymes, but it appears that foregut fermentors such as ruminants have evolved lysozymes produced in the abomasum as part of their digestive strategies. There is also evidence that rabbits have also evolved a similar mechanism, but in this case the lysozyme is secreted by the colon wall as the membrane is secreted onto the soft pellet.
Lysozymes are a group of enzymes that specifically digest the mucopolysaccharide, peptidoglycan of bacterial cell walls. The peptidoglycan envelop appears to slow the potential digestion of bacterial cells and the short small intestine of the rabbit may limit efficiency of digestion of caecal microbes. It is hypothesised here that lysozyme is involved in the digestion of bacteria in the caecotropes in the stomach.
To enhance the digestion of bacteria it is also hypothesised that bacteriophage action is also involved in lysis of bacteria in both the stomach of the rabbit and the abomasum of the ruminant.
The production of caecotropes, with their high protein content also distributes protein digestion and amino acid availability over a prolonged period of time. A process that maybe necessary because of the apparent relatively low digestion of protein that has been shown by studies on the ileal flow of protein on high protein diets. High protein in a diet seems to allow a detrimental microbial population to develop in the lower digestive tract of the rabbit leading to high death rates.
The efficient use of forage based diets, and capacity to breed with multiple offspring, demarcates a special role for the forage-fed rabbit when oil prices rationalise the use of grain for intensive animal production
Rabbits are herbivores and are classified as hindgut (caecum and colon) fermentors. In nature they are highly selective feeders and they can efficiently digest a wide range of simple and complex carbohydrates by curtsey of their digestive strategy. The rabbit has an efficient monogastric mode of digestion that is followed by fermentation of ‘selected’ cellulose feed and endogenous materials in the caecum through the action of a resident bacterial ecosystem comprised primarily of Bacteroides spp
Because of a small body size [large surface area to weight] the rabbit has a high metabolic rate and therefore a relatively high maintenance energy requirement particularly when compared to ruminants and equines. Forage intake in the former is constrained by rate of comminuting feed to small particles in the rumen: The horse on the other hand compensates for low digestibility of forage by eating more feed and passing this through the gut rapidly; sacrificing digestibility to meet energy demand. The pig is limited in its use of forage by the slow movement of cellulosic materials through the large intestine which feed back to limit intake.
On concentrate diets, feed intake in the rabbit has to be in excess of 5% of live weight per day (Irlbeckel 2001) and as high as 8-10% of live weight per day on forage based diets (Pok Samkol et al 2006b) to ensure nutrient availabilities are above maintenance. These levels of intake are achieved through a special adaptation of digestive processes that allows the rabbit to efficiently digest non-fibrous carbohydrates, but quickly exclude from its digestive tract the relatively indigestible fiber in a diet and at the same time preserve digestion of readily fermented fiber [polysaccharides] in the caecum.
The rabbit caecum is very large, compared with the rest of the gut (Stevens and Hume 1995) and forms a spiral that fills the abdominal cavity (see Figure 1).
|
|
|
Figure 1: The digestive tract of the rabbit |
The caecum has a capacity 10 times that of the stomach, and occupies about 40% of the gastrointestinal tract in concentrate fed rabbit but this may be greater in forage fed animal (Jenkins 1999). Rabbits utilize a mechanism to expel slowly fermented fiber from the gastrointestinal tract, whilst retaining readily fermented materials and endogenous nitrogenous components of the digesta leaving the ileum. Consumption of soft faeces [caecotropes] produced from the materials that pass through the caecal cycle also compensates for low or poor-quality protein in a diet by recycling, via fermentation in the caecum, endogenous protein secretions and sloughed epithelial cells from the small intestine (Carabano and Piquer 1998).
Feed undergoes enzymatic digestion in both the stomach and small intestine as the digesta moves through the tract. The small intestine in the rabbit is relatively short so only readily digestible materials are extracted such as sugars, soluble protein and particulate starch. Adult rabbits appear to have little capacity to digest fat in the small intestine and lipase activity is restricted to the caecum (Marounek et al 1995). High amounts of fat in a diet may therefore limit cellulose fermentation in the caecum as shown for the forestomach of ruminants (see Devendra and Lewis 1974). Fat deposition on fiber, increasing its density, may also be a factor in allowing the separation of the course fibers away from the caecum, and their subsequent excretion as components of the hard faeces.
When digesta from the ileum enters the large intestine, muscular contractions facilitate the separation of slowly digestible fiber from other materials including non fiber fractions (eg: protein and soluble carbohydrates) and small fibrous components. A series of strong peristaltic contractions move fibrous particles through the colon towards the anus and anti peristaltic waves move liquid and small particles back into the caecum. In this way the rabbit separates and concentrates digesta fractions which are readily degradable for preferential fermentation in the caecum ( Lebas et al 1997; Carabano and Piquer 1998); particle size and fluid density aid separation (Cheeke 1994). The relatively indigestible fiber components that separate out are voided quickly as hard faeces (Figure 2) about 4 h after consumption of a meal (Cheeke 1994).
|
|
|
Figure 2: Caecotropes (left of picture) from the fundus compartment of the stomach and hard faeces (right of picture) |
Fermentation of the materials that are directed into the caecum, proceeds for some further 4 hours. During this time, contractions of the caecum move the lumen digesta contents towards the blind sac and back and at the same time mixes the contents. Fermentation of solubles and fibrous materials is likely to be efficient in terms of microbial growth since the residence time of digesta is only 4h compared with retention times of feed in the rumen often greater then 12h.
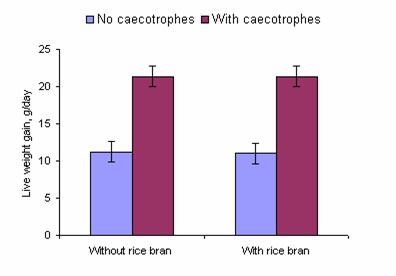
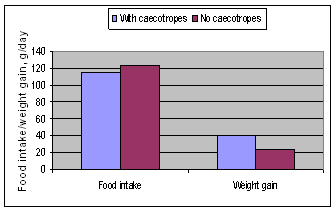
Small discrete digesta samples are periodically forced by muscular contractions into the large intestines where some moisture is absorbed along with some [most?] VFA (Stevens and Hume 1995). As the digesta from the caecum is propelled towards the anus it forms into pellets which are coated with a mucous membrane secreted by the cells of the colon wall to become caecotropes. The caecotrope is voided from the body approximately 8 hours after consumption of a meal (Cheeke 1994). The rabbit recognizes the caecotrope to be voided and consumes it directly from the anus. This practice of consuming caecotropes is called coprophagia, or caecotrophy. Under natural or grazing conditions, caecotrophy usually occurs during the day and feeding is restricted to the night time. Hard faeces and soft faeces are excreted in a circadian rhythmic pattern (Carabano and Piquer 1998; Lebas et al 1997). If a rabbit is equipped with a collar preventing caectrophy, the digestion of the diet is significantly reduced and growth rates are often decreased by some 50% (Figure 3a and b).
|
|
|
Figure 3a: Effect of access or not to caecotropes on the growth rate of rabbits fed water spinach with or without supplementation with rice bran (Chiev Phiny and Lampheuy 2006) |
|
|
|
Figure 3b. Food intake and weight gain in New Zealand White rabbits fed a concentrate pellet (Stephens 1977) |
The caecotropes concentrate considerable valuable nutrients that the rabbit uses . A comparison of nutrient content of hard faeces v caecotropes is shown in Table 1. When allowed to select their diet, rabbits will select the least lignified, succulent plant parts or the plant parts that are most nutrients dense and highest in sugar. This feeding strategy allows the animal to meet the dietary requirements for their high metabolic rate (Cheeke 1994). Rabbits eliminate relatively indigestible fiber as quickly as possible from their gastrointestinal tract.In many situations this would be the most lignified fibrous materials. Rabbits have high feed intake and fast feed transit time, which enable them to consume forages and meet nutritional requirements (see Pok Samkol et al 2006b; Carabano and Piquer1998). Understanding the digestive physiology of the rabbit and how dietary components affect microbial growth efficiency in the caecum is a key to proper feeding management.
|
Table 1. Nutrient composition of caecotropes and hard faeces (Carabano and Piquer 2003) |
||
|
Material |
Caecotropes |
Faeces |
|
Dry Matter, % |
34 |
47 |
|
Protein, % |
30 |
17 |
|
Crude Fiber, % |
18 |
30 |
|
Potassium, mmol/kg DM |
260 |
84 |
|
Bacteria 1010/g DM |
142 |
31 |
|
Nicotinic acid, mg/kg |
139 |
40 |
|
|
|
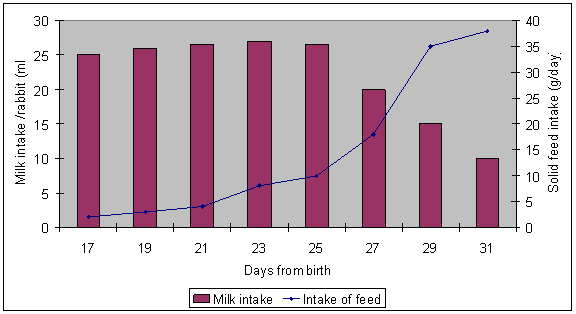
Figure 4. Relationship between milk intake by young rabbit and solid feed intake up to weaning ( 10 kits/litter) (Fortun-Lamothe and Gidenne 2000) |
The caecum has specialized properties that allow rabbits to be fed a wide range of feeds. In the young rabbit, whilst suckling, the caecum remains relatively free of microorganisms until the commencement of solid food intake. During the suckling period the young rabbit depends largely on protein and fat in the mother’s milk which has very low lactose content (around 1% of milk). Solid feed intake begins about 17 days and gradually increases until weaning at about 1 month of age (see Figure 4. Fortun-Lamothe and Gidenne 2000)
The volume of the caecum and of the colon increases linearly to become the largest organ in the digestive tract in 5-6 weeks from birth. The dry matter content of the caecum /colon comprises about 40% of the total digesta volume and has a dry matter content of about 22%. The microbial species present are influenced extensively by the diet particularly when solid feed intake is high-just prior to weaning. Milk has a major bacteriostatic role as it contains a high milk fat that is high in C8 and C10 fatty acids which suppresses microbial colonization of the anterior part of the tract (Cole et al 1982).
As milk consumption decreases with age, there is a concomitant increase in the intestinal flora. The caecum /colon harbour an abundant flora from the first week. The facultative anaerobic flora is dominated largely by Streptococcus species until 14 days of age. A mixed population of microbes exist in the digestive tract mainly dominated be Bacteroides species but also present are sporulating bacteria such as Clostridium, Endospous and Acuformis which if they suddenly receive high starch intake as the young rabbit commence to take in solid feed, can become dominant with losses due to the effects of the resultant bacteria . The bacteria involved in fermentation of fibrous carbohydrates only become dominant after 15 days of age, coincident with intake of fiber. Rabbits receiving only milk, have no cellulolytic flora (Padilha et al 1999) and it is presumed that rabbits given starch will develop a totally different flora. Sometimes leading to diahorea .
Surprisingly rabbits that consume high protein diets are more prone to die both immediately following weaning and during the whole fattening period. ( see Chamorro et al 2007) ( see Table )which appears to be associate with more protein reaching the lower tract and proliferation of Clostridia and Escheria Coli (Cotez et al 1992) The ideal diet for the young rabbit just prior to weaning is a low protein high fiber in which the fiber contains high levels of pectins and xylans as the organisms that digest these are the most viable in the early stages of the life of the rabbit ( see Gidenne and Fortun-Lamothe 2002.
It appears that the protein in a high protein diet is incompletely digested in the short intestine and when the enzyme system is over whelmed, protein enters the lower tract causing a detrimental change in the flora which leads to a high death rate in rabbits.
The supplementation of low protein diets with microbial protein produced in the caecum and returned to the gut via caecotropes may have evolved to spread the availability of protein for digestion and prevent excess protein entering the lower gut at any time since it is held for a considerable time in the stomach.
|
Table 2. The effects of different levels of protein in four iso-energetic and iso-fibrous diets on mortality of rabbits shortly after weaning and over the fattening period (Chamorro et al 2007). Lucerne hay was substituted by other fibrous feeds (largely wheat straw) and soybean protein replaced pulps (largely apple and beet). |
||||
|
Diets |
High protein/high lucerne |
Low protein/high lucerne |
Low protein/ medium lucerne |
Low protein/low lucerne |
|
Mortality-14 days post- weaning plus added antibiotics |
7.7 |
1.9 |
1.9 |
0 |
|
Mortality -14 days post weaning (%) |
33 |
20 |
23 |
20 |
|
Mortality whole fattening period plus added antibiotics |
9.6 |
1.9 |
1.9 |
3.8 |
|
Mortality whole fattening period (%) |
33 |
20 |
24 |
23 |
It is particularly difficult to prevent the young rabbits with their mothers from consuming the mother’s diet, which should be designed to provide a relatively high energy-high protein to support substantial milk production. However to prepare the young rabbit for weaning the diet required should be relatively low in protein and high in structural carbohydrates to adapt the caecal organisms and avoid the problems of diarrhea associated with the weaning period . Where does are fed complex diets in late gestation it might be appropriate to either find a way to provide a more fibrous diet low in true protein for the young animals or to revert the doe to this diet for 10 days prior to weaning.
Based on the production records of around 850 rabbit farms (Guerder 2001) about 30% of all young rabbits die between birth and slaughter in the industrialized feeding systems used for example in France. This is on top of high losses (20 to 30%) and a short
reproductive life (<10 months) for the doe. Surveys of the losses in non industrialized countries developing rabbit farms do not appear to exist. However they may not be so high because of the lower levels of concentrates used in these systems.
One criticism aimed at the systems of forage-fed rabbits is that growth rate is approximately half that in the concentrate-fed rabbit. However, the losses from early deaths of young rabbits and does alike probably offset the lower growth rate. A research programme to look at life time productivity on the two systems appears to be warranted.
I t appears that survival of young animals and longevity of breeding stock is highly associated with successful production of caecotropes in the young animal as it becomes dependent of solid feed and the older animal as the caecal health applies to the type of feed provided.
When compared with other herbivores, digestion of structural components of plants such as alfalfa in mixed concentrate based diets for rabbits is relatively low (14% for alfalfa hay in rabbits compared with 44% in cattle, 41% in horses, and 22% in pigs) (McNitt et al 1996). The actual crude fiber component of most forages fed to rabbits in industrialized countries is only 20 to 25% (McNitt et al 1996), depending on forage source and its maturity. Other non fiber fractions of forage such as protein, and soluble carbohydrates are easily digested by rabbits. In concentrate-fed rabbits supplemented with forages such as lucerne the dietary fiber is structural and often closely associated with lignin and has a critical role in maintaining gut health, stimulating gut motility (insoluble fiber only), reducing fur chewing, and preventing enteritis (McNitt et al 1996; Brooks 1997). Concentrate-based diets with less than 20 to 25% fiber result in reduced gut motility, reduced caecotrope formation, prolonged retention time in the hindgut, and often enteritis and diarrhea (Cheeke 1994; Jenkins 1999).
In the forage-fed rabbit the diet is a mixture of soluble sugars, protein and fiber that may be relatively un-lignified or highly lignified according to source. For instance the rabbit fed water spinach or tree leaves consumes little lignified fiber. In these diets it is the soluble components and readily fermented structural carbohydrates that provide the energy and protein requirements .On the other hand when fed grasses or tall growing legumes the fiber can be highly lignified as it supports the upright growth of the plant.
Composition of the hard feces and the caecotrope is influenced by the diet. If dietary fiber concentration increases, the fiber composition of the faecal pellets also increases. Fiber fermentation in concentrate-fed rabbits does not seem to be enhanced by coprophagia (as cited by Cheeke 1994) but it is clearly influenced when rabbits receive only forage as the basis of the diets (Chiev Phiny and Lampheuy 2006).
Fermentation in the caecum results in an increase in microbial biomass and the production of volatile fatty acids [VFA] in a similar way as occurs in the foregut fermentation of ruminants. However there are potentially significant differences in the efficiency of net microbial growth in the rabbit caecum as compared with the ruminant fore stomach in both the forage and concentrate fed animal
In rabbits fed a traditional alfalfa/maize diet, acetate is the primary volatile fatty acid (VFA) produced by microbes, with more butyrate than propionate being formed. Microbes in the rabbit’s caecum produce more VFA on starch-based diets than on forage diets (Cheeke 1994), which may be linked with the efficiency of microbial growth rather then differences in total dry matter digestion by fermentation. Stevens and Hume (1995) indicate that VFA provide a major energy source in the rabbit colon.
Even though amino acids from bacterial protein may be available via coprophagia (especially lysine, sulfur amino acids, and threonine; Carabano and Piquer 1998), research has shown that microbial protein plays only a minor role in meeting a rabbit’s protein and amino acid needs on diets based on concentrates (McNitt et al 1996). The majority of microbial protein utilized by the animal is digested in the colon (Stevens and Hume 1995), presumably once the caecotrope disintegrates and its contents are released in the stomach. Caecotropes, however, contain approximately 28-30% crude protein (Stevens and Hume 1995 see Table 1) which may indicate they are approximately 50% bacterial cells as bacterial cells from the rumen are 50-60% crude protein. The low apparent utilization of microbial protein may be caused by the diet. Recent studies with rabbits on forage-based diets indicate that prevention of caecotrope consumption has big effects on growth of rabbits, which is not in line with the concept that they supply little extra nutrients as VFA and essential amino acids of microbial origin. .
Rabbits can digest 75 to 85% of lucerne protein, whereas pigs digest less than 50% (McNitt et al 1996).Urea is recycled by the rabbit large intestine in a manner similar to that occurring in the rumen (Stevens and Hume 1995) and urease levels are highest in the caecum (Marounek et al 1995). Urea is converted to ammonia in the gut, and if the content of carbohydrate is low, the ammonia may be absorbed and this could result in toxicity. When an animal is fed a low-energy diet, caecotrope ingestion is maximized (Jenkins 1999). Low levels of dietary protein fed to rabbits increase caecotrope consumption and high levels of protein decrease consumption, which seems to be a protein sparing mechanism (Cheeke 1994). Coprophagia has been found to increase protein digestibility (50 vs 75 to 80% for alfalfa) of forages in rabbits
In any fermentative system VFA production and microbial growth are linked; the energy for microbial growth is provided by ATP produced when carbohydrate and protein are degraded to VFA. The microbial polymers or building blocks for microbial cells are synthesized from the intermediates of glycolysis and VFA formation (Preston and Leng 1987). Y-atp (Bauchop and Elsden 1960) is a useful description of the efficiency of microbial growth. It is defined as the g dry cells produced in fermentation per mole of ATP available in the conversion of organic matter to VFA
In the rumen Yatp is much reduced by the long retention time of feed particles together with a high maintenance requirement of the microbes. In addition a considerable amount of lysis of microbes occurs from the action of lytic phage, autolysis and predation by the protozoa (Wells and Russell 1996; Klieve and Swain 1993; Leng and Nolan 1984). Predation of protozoa can be significant (Bird and Leng 1984) often decreasing the net availability of microbial protein by 25-35% (Bird and Leng 1984) but, in addition, protozoa have a high maintenance energy requirement (use ATP for maintenance rather then growth) and are retained in the rumen for longer periods than indicated by rumen turnover time (Weller and Pilgrim 1974). Most measurements for Yatp in the rumen suggest a value of 8-14 g cells/mole of ATP generated in fermentation, whereas the theoretical Yatp should be about 26 g cells per mole of ATP. The Yatp for fermentation in the caecum of rabbits, to my knowledge has not been measured, but there are a number of indications that many of the inefficiencies in the rumen do not apply to the rabbit’s caecal environment. These include i) the absence of protozoa; ii) the absence of slowly fermented fiber which has been rejected through the sieving mechanism in the large intestine; iii) the short turnover time of the caecum’s contents; iv) the removal of anti bacterial components in the upper digestive tract; v) the enrichment of the medium by secretions from the small intestine including secreted proteins such as enzymes, mucous secretions and sloughed cells of intestinal origin; and vi) the small number of bacterial species which will much reduce inter-species feeding.
With these advantages it is feasible that the end products of fermentation in the rabbit caecum will be richer in cells and therefore in protein (bacteria are 50-60% protein) than is the case for rumen fermentation. This potentially rich source of essential amino acids is then made available to the rabbit through coprophagia. This idea is supported by the very high protein content of the caecotrope (28% CP in DM see Stevens and Hume(1995)and 47% in DM in rabbits fed water spinach leaves or stems as the sole diet (Pok Samkol et al 2006a).
A high efficiency of microbial growth depends to a major extent on a rapid turnover of caecal contents in rabbits reducing bacterial lysis and bacterial maintenance energy requirements. In the pig given high fiber diets the bacterial protein from the large intestine is unavailable to the animal and fermentation is a source of energy substrates in the form of VFA. A slow turnover of the contents of the large intestine is therefore beneficial if the pig is on a high fiber diet as lysis of cells and fermentation of bacterial proteins allows more VFA absorption (Marounek et al 2002) As shown by Dierick et al (1990) the bacterial growth efficiency in the pigs’s hind gut is less then half that in the rumen (13.6 g bacterial N excreted in the faeces per kg of organic matter apparently fermented compared to 30 g N/kg organic matter fermented in the rumen). The latter is approximately a third of the theoretical efficiency of anaerobic bacterial growth.
The sieving and rejection of fibrous feed from entry into the caecum may have further implications for bacterial growth efficiency in the caecum. As the largest, intact, fibrous components in digesta entering the large intestine are likely to be the most lignified, the “sieving” action in the caecum/colon may remove these preferentially and decrease the lignin content of organic matter in the caecum relative to that in fiber in the diet or in the hard faeces. The significance of this is that the release of phenolic compounds from lignin has been shown to suppress bacterial growth in the rumen (see Borneman et al 1986) and reduce digestibility of both starch and cellulose. For example whole oats and oat hulls with high lignin content often have a much lower digestibility then varieties that have low lignin content (see Black 2001: Rowe and Crosbie 1998). Thus a physiological mechanism excluding lignin from the caecum could potentially improve the efficiency of bacterial growth. There is a remarkable capacity for degrading pectins and hemicelluloses, attributable to the substantial establishment of xylenolytic flora compared with the cellulolytic flora (see. Gidenne and Fortun-Lapmothe 2002)
The caecotrope which is taken from the anus directly and swallowed remains in the stomach for some hours. The tough membrane remains intact for at least six hours after ingestion. When swallowed the caecotropes pass to the fundus portion of the stomach (Griffiths and Davis 1963) which is at almost neutral pH The membranes around the pellet and a buffering solution in the pellet control pH and fermentation seems to proceed even though the rest of the stomach is acid Cheeke (1994) has suggested that the VFA production is not significant but the permeability of epithelial cells to VFA is high and so the VFA are probably mostly absorbed from the caecum, large intestine and colon and at high efficiencies of growth are in much lower proportions in the end products [cells and VFA]. The purpose of the caecotrope appears to be to allow a prolongation of fermentation activity while the caecotrope is resident in the stomach. When feeding commences and these disintegrate they make available bacterial cells and micronutrient to be used in intestinal digestion.
Is there another reason for the residence of the caecotrope in the stomach? The movement of digesta materials through the stomach- small intestine is quick and the small intestine is short suggesting that digestion of bacterial cells which are enclosed in a highly resistant membrane envelop of peptidoglycan, may be too slow allow significant significant digestion.
The possibility is floated here that caecotrophy may allow an efficient use of bacterial cellular materials. In the process of formation of the caecotrope and its retention in the stomach, activation of temperate phage to its lytic phase in bacterial cells causes bacteria to lyse improving the digestibility of bacterial protein in the stomach and small intestine. Protein may also be partially digested by enzymes released during lysis.
In ruminants lysozyme produced in the abomasum assists in the degradation of the ruminants major essential amino acid supply - the microbes produced in the rumen.
Lysozymes are a group of enzymes [muramidases] that hydrolyses beta-1,4-links between N-acetyl-muramic acid and N-acetyl-D-glucosamine in the peptidoglycan of bacterial cell walls .The enzyme is found in tears, saliva, white blood cells and macrophages where it may have a defense role against invasive organisms.
A group of lysozymes appear to have a digestive role in the true stomach (abomasum) of ruminants and colobine monkeys and also copraphagic animals. These enzymes degrade the mucopolysaccharide cell walls of bacteria passing from the rumen, allowing the cell contents to be digested by other stomach and intestinal enzymes (Dobson et al 1984; Stewart et al 1987; Irwin and Wilson 1989). The stomach form of lysozyme is endowed with special physiochemical properties that allow it to function in an acidic and protease-rich environment.
In the rabbit, lysozyme produced in the non-sacculated colon is secreted circadially into the colonic lumen in association with the production of caecotropes that are destained for ingestion (Camara and Prieur 1984). Because of the mucous coating of the caecotrope, the lysozymes are concentrated in the caecotrope. Thus from the time the digesta leaves the caecum and forms a caecotrope and is then excreted and swallowed, to the time it disintegrates in the stomach there is a considerable opportunity for lysozyme to degrade the bacterial cell peptidoglycan. However, the lysozymes appear to have narrow and acidic pH optima (Ito et al 1994), and therefore they are more active in the low pH of the stomach. The action of lysozyme could be considerably enhanced if phage-initiated disruption of the bacterial cell membrane occurred prior to, following or coinciding with the change in pH as the caecotrope disintegrates in the stomach
Another interesting fact is that bacteriophages also contain lysozymes which appear to have a pH optima close to neutral. Phages infect cells by attaching to the cell via a base plate and forcing their tail tube into the cell membrane puncturing the outer membrane. Contractions of the sheath progresses enlarging the pore in the membrane until the lysozyme domains reach the peptidoglycan layer which they digest (see for more detail Kanamaru et al 2002). The possibility exists that bacteriophage activity is stimulated in the close to neutral caecotrope prior to the onset of colonic lysozyme activity when activated by the acid environment of the stomach,
Large numbers of temperate and lytic phage are associated with rumen organisms (Orpin and Mann 1974: Klieve and Swain 1993). We have no knowledge of their presence in caecal fluid organisms but it appears logical that the phages are present in these organisms since the major bacteria present in caecal contents [Bacteriodes species] are known to be infected with phage in the rumen (Keller and Traub 1974).
As an aside to the rabbit, and pertinent to the ruminant, it appears possible that rumen organisms entering the abomasum would be killed by the acid environment, and for the bacterial cell envelope of peptidoglycan to remain intact, temperate phage would quickly multiply and become lytic phage and assist the lysozymes of the acid abomasum to complete the disruption of the bacterial cells and enhance the process of their digestion. It thus seems that similar systems to degrade bacteria have evolved in both foregut and hind gut fermentors via caecotrophy in the latter.
In summary, in the rabbit the lysozyme that is produce by the colon cell wall but conserved in the caecotrope may play a major role in facilitating the digestion of bacteria in the stomach and small intestine. Elucidating the potential role of phages as aides to bacterial cell digestion needs considerable research
Efficient fermentation (high cell yield relative to VFA production in the caecum) and efficient digestion of bacterial cells by combined action of lysozyme, potentially lytic phage and gastric and intestinal enzymes, may account for a very superior mode of both conserving and utilizing essential amino acids by the rabbit.
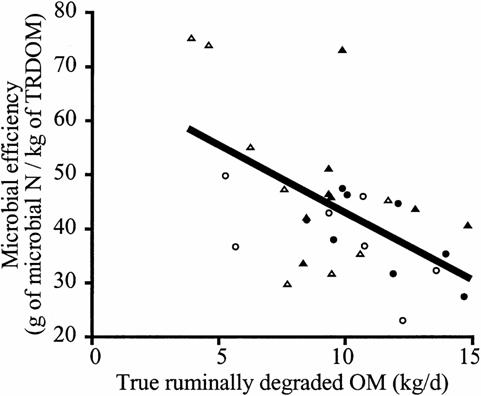
High-starch diets are often incompletely digested in the rabbit small intestine due to rapid transit times (McNitt et al 1996). Incomplete digestion of starch prior to the large intestine results in the availability of starch for microbial fermentation (Stevens and Hume 1995). Excess starch in the gut results in an extremely rapid fermentation with possible spilling of energy leading to low Yatp. For example, Oba and Allen (2003) showed that, as the rate of starch digestion increased, the Yatp was reduced from 60g of microbial N/kg starch digested to approximately 30 g microbial N/ kg starch digested (see Figure 4).
|
|
|
Figure 4. Relationship between rate of starch digestion in the rumen of cows and microbial efficiency.. Closed circle denotes high moisture maize in high starch diets; closed triangle denotes dry ground maize in high starch diets; open circle denotes high moisture maize in low starch diets; and open triangle denotes dry ground maize in low starch diets (after Oba and Allen 2003) . |
The rapid digestion of starch could thus lead to a much lower microbial cell yield in starch fermentation in the caecum of concentrate-fed as compared to forage-fed rabbits. If toxin-producing microbes (primarily Clostridium spiroforme) are in residence, high levels of starch may lead to enteritis and possible death (McNitt et al1996; Jenkins 1999).
Grains processed too finely can lead to rapid bacterial fermentation of the starch and cause enterotoxaemia. Rabbits were never meant to be fed high starch diets; and on high forage diets, with little complex structural carbohydrates, their feeding strategies, combined with their digestive strategies, could be much more of an advantage than when fed diets based on lignified forage, cereal grains and protein meals.
The use of maize-alfalfa diets for feeding rabbits is an expensive option, that has developed because of the economic advantages of industrial production systems in countries with high labour costs, such as the USA and in the EU. Such feeding systems are designed along similar lines as for other monogastric animals and may remove the significant advantage of the rabbit - the ability to utilize caecotrophy as a tool to optimize essential amino acid availability. The feeding of high-starch diets to rabbits may lead to reduced microbial growth efficiency in the caecum leading to insignificant improvements in protein nutrition via caecotrophy. In contrast, on high-forage diets this process has much more significance if, as suggested in this review, microbial growth in the caecum is highly efficient.
Feeding a traditional alfalfa and maize diet to rabbits is not likely to be desirable nor economic in the future as resource depletion and competition for grain for feed, food and feedstock, forces up the price of grain (Preston 2006).
Ultimately, efficient feeding of rabbits should be based on locally available forages of high nutritional value, such as herbs and tree leaves and specialized high yielding crops such as water spinach. The moderate protein content of such feeds can then be enhanced by the rabbit’s evolved digestive strategy for using forages. This feeding strategy maintains healthy microbial digestion in the caecum and optimizes the potential advantages of rabbits over other herbivores. When combined with their capacity to produce multiple offspring at 6 week intervals, rabbits have a tremendous capacity to produce animal protein for human consumption without competing for fossil fuel costly feed.
The potential feed sources for rabbits are widely available in most tropical countries in Asia. However, any feeding system should recognize that the time of supplying feed should not disturb the potential circadian rhythm of caecotrope production. Considerable research is needed with fiber based diets to find the optimum pattern of offering feed that will enhance the production and utilization of caecotropes by the animal. Furthermore, the supply of small amounts of starch-based feed as a single meal early in the day [a common practice in many tropical countries] may interrupt the residence time of caecotropes in the stomach and may also cause disruption of the fermentative efficiency in the caecum, particularly if the feed contains appreciable enzyme-resistant starch mainly amylose.
Brooks D 1997 Nutrition and Gastrointestinal Physiology. In: E V Hillyer and K E Quesenberry (ed.) Ferrets, Rabbits and Rodents— Clinical Medicine and Surgery. p 169. W.B. Saunders Company, Philadelphia.
Bauchop T and Elsden S R 1960 The growth of microorganisms in relation to their energy supply. Journal of General Microbiology 23:457
Bird S H and Leng R A 1984 Further studies on the effects of of the presence or absenceof protozoa in the rumen on the live weight gain and wool growth of sheep. British Journal of Nutrition 52 607
Black J L 2001 Quality Feed Grains – Research highlights and opportunities. Proceedings of the 10th Australian Barley Technical Symposium http://www.regional.org.au/au/abts/2001/m3/black.htm retrieved 19th Oct 2006
Borneman, W S Akin D E and van Eseltine W P1986. Effect of phenolic monomers on ruminant bacteria. Appl. Environ. Microbiol.52:1331–1339.
Camara V M and Prieur D J 1984 Secretion of colonic isozyme of lysozyme in association with caecotrophy of rabbits. American Journal of Physiology Gastrointestinal and Liver Physiology 247, G19-G23
Carabano R and J Piquer 1998 The Digestive System of the Rabbit. In: C de Blas and J Wiseman (ed.) The Nutrition of the Rabbit. p 1. CABI Publishing, London.
Carabano, R., and J. Piquer. 2003. The digestive system of the rabbit. In: The Nutrition of the Rabbit. Edd C. de Blas and J. Wiseman. CABI Publishing, Oxon, UK pp 1-16.
Chamorro S., Gomez-Conde M. S.,. Perez de Rozas A. M, Badiola, , I. Carabanoand R. De Blas J. C. (2007) Effect on digestion and performance of dietary protein content and of increased substitution of lucerne hay with soya-bean protein concentrate in starter diets for young rabbits Animal 1;651-659
Cheeke P R 1994 Nutrition and Nutritional Diseases. In: P J Manning, D H Ringler and C E Newcomer (ed.) The Biology of the Laboratory Rabbit. 2nd ed. p 321. Academic Press, New York.
Chiv Phiny and Lampheuy Kaensombath 2006: Effect on feed intake and growth of depriving rabbits access to soft faeces. Livestock Research for Rural Development. Volume 18, Article # 34. Retrieved, from http://www.cipav.org.co/lrrd/lrrd18/3/phin18034.htm
Cole C B, Scott K J, Henschel M J, Coates M E, Ford J E and Fuller R 1982. Trace nutrient binding proteins in milk and the growth of bacteria in the gut of infant rabbits. British Journal of Nutrition 49: 231-240.
Dierick N A, Vervaeke I J, Decuypere J A, Henderickx H K 1990 Bacterial protein synthesis in relation to organic matter digestion in the hindgut of growing pigs; contribution of hindgut fermentation to total energy supply and growth performances. Journal Animal Physiology and Animal Nutrition 63:,220-235
Devendra C and Lewis D 1974 The interaction between dietary lipids and fibre in the sheep. Animal Production (19), 67-76.
Dobson D E, Prager E M and Wison A C 1984 Stomsach lysozymes of ruminants. 1 Distribution and catalytic properties The Journal of Biological Chemistry 259,11607-11616
Fortun-Lamothe L and Gidenne T 2000. Effects of the suckled litter size on intake behaviour, performance and health status of young and reproducing rabbits. Annales de Zootechnie 49: 517-529.
Gidenne T and Fortun-Lamothe L 2002. Feeding strategy for young rabbits around weaning: a review of digestive capacity and nutritional needs Animal Science 75 169-184
Guerder F 2001. Résultats de GTE Renalap. Cuniculture 28: 171-175.
Griffiths M and Davies D 1963 The role of the soft pellets in the production of lactic acid in the rabbit stomach .Journal of Nutrition 80,171-189
Jenkins J R 1999 Feeding Recommendations for the House Rabbit. Veterinary Clinics of North America: Exotic Animal Practice. vol. 2. p 143. W.B. Saunders Company, Philadelphia
Irwin D M and Wilson A C 1989 Multiple cDNA sequences and the evolution of bovine stomach lysozyme. Journal of Biological Chemistry 264(19):11387-11393
Ito Y, Hirashima M Yamada H and Imoto T1994 Colonic lysozymes of rabbit [Japanese White]:Recent divergence and functional conversion. Journal of Biochemistry 116, 1346-1353
Kanamura S, Lehman PG, Kostyuchenke V A,Chipman P R, Mesyanzhinov V V, Arisaka F and Rossman M G 2002 Structure of the cell-puncturing device of bacteriophage T4 Nature 415, 553-557
Klieve A V and Swain R A 1993 Estimation of ruminal bacteriophage numbers by pulsed-field gel electrophoresis and laser densitometry. Appl Environ Microbiol. 1993 July; 59(7): 2299–2303
Lebas F, Coudert P, Rochambeau H de and Thébault R G 1997 The Rabbit - Husbandry, Health and Production. FAO Animal Production and Health Series No. 21 http://www.fao.org/docrep/t1690E/t1690E00.htm
Leng R A and Nolan J V 1984 Nitrogren metabolism in the rumen. Journal Dairy Science 67 1072- 10??
Marounek M, Adamec T , Skfiivanova V and Latsik N I 2002 Fractions of Nitrogen and in vitro Fermentation of Nitrogenous Substrates in Caecal Contents of the Pig. Acta Vet. Brno 2002, 71: 429-433.
Marounek M,Vovk S J and Skfiivanova V(1995) Distribution of activity of hydrolytic enzymes in the digestive tract of rabbits. British Journal of Nutrition 73,463-469
McNitt J I, Cheeke P R, Patton N M and Lukefahr S D 1996 Rabbit Production. Interstate Publishers, Inc., Danville, IL.
Oba M and Allan M S 2003 Effects of diet fermentability on efficiency of microbial N production in lactating dairy cows J Dairy Science 86 195 -207
Orpin C G and Mann E A 1974 The occurrence of bacteriophage in the rumen and their influence on the bacterial population Experientia 30, 1018-1020
Padilha, M T S, Licois D, Gidenne T and Carré B 1999. Caecal microflora and fermentation pattern in exclusively milk-fed young rabbits. Reproduction, Nutrition, Development 39: 223-230.
Preston T R 2006: Forages as protein sources for pigs in the tropics. Workshop-seminar "Forages for Pigs and Rabbits" MEKARN-CelAgrid, Phnom Penh, Cambodia, 22-24 August, 2006. Retrieved, from http://www.mekarn.org/proprf/preston.htm
Pok Samkol, Preston T R and Leng R A 2006a Effect of offering leaves or stems of water spinach on patterns of eating, consumption of caecotrophs, and excretion of faeces by growing rabbits. Livestock Research for Rural Development. Volume 18, Article # 78. RetrievedOctober 16, 106, from http://www.cipav.org.co/lrrd/lrrd18/06/samk18078.htm
Pok Samkol, Preston T R and Ly J 2006b Effect of increasing offer level of water spinach (Ipomoea aquatica) on intake, growth and digestibility coefficients of rabbits. Livestock Research for Rural Development. Volume 18, Article #25. Retrieved March 7, 2006, from http://www.cipav.org.co/lrrd/lrrd18/2/samk18025.htm
Rowe J B and Crosbie G B 1998 The digestibility of grains of two oat s differing in lignin content Australian J Agricultural Research 39 639-644
Stewart C B, Schilling J W and Wilson A C 1987 Adaptive evolution in the stomach lysozymes of foregut fermenters. Nature 330(6146):401-404
Stephens 1977
Digestibility and coprophagy in the growing rabbit. Proceedings of the Nutrition
Society 36, 4A
Stevens C E and Hume I D 1995
Comparative Physiology of the Vertebrate
Digestive System. 2nd ed. Cambridge University Press, Cambridge, United Kingdom.
Keller R and Traub N 1974 The characterization of Bacteroides fragilis bacteriophage recovered from animal sera: observations on the nature of bacteroides phage carrier cultures. Journal General Virology 24: 179 - 189.
Weller and Pilgrim 1974 Passage of protozoa and volatile fatty acids from the rumen of sheep and from a continuous in vitro fermentation system. British Journal of Nutrition 323, 341 -351
Wells J E and Russell J B 1996 Why do so many ruminal bacteria die and lyse so quickly. Journal of Dairy Science 79 1487-1495