| Cassava leaves for monogastric animals |
Nutritive value of cassava
leaves for monogastric animals
Bui
Huy Nhu Phuc, Brian Ogle* and Jan Erik
Lindberg*
University
of Agriculture and Forestry,
Thu Duc, Ho Chi
Minh City, Vietnam
* Swedish University of Agricultural
Sciences,
PO Box 7024, SE-750
07 Uppsala, Sweden.
Abstract
Leaves from varieties of cassava (Manihot esculenta Crantz) growing in
South Vietnam were collected at the time of harvesting the roots and analysed
for their nutrient and cyanide content. Nutritional properties of the dried and
ensiled cassava leaves were determined with pigs and rats.
The crude protein of cassava leaves
ranged from 240 to 350 g/kg, and the crude fibre from 100 to 150 g/kg on a dry
matter basis. It was found that the HCN content was markedly reduced by both
drying and ensiling, with the reduction being highest after sun-drying and
oven-drying. The ileal and total tract apparent digestibilities in pig and rats
of organic matter, crude protein, nitrogen free extractives and energy, and
nitrogen retention, were negatively affected by inclusion of cassava leaves. The
impaired amino acid profile after drying cassava leaves at 105 oC was
found to be related to a slight decrease in digestible protein as well as N
retention. Average ileal
digestibility coefficients of organic matter, crude protein and crude fibre in
cassava leaves were 0.42, 0.37 and 0.35; total tract digestibility coefficients
were 0.57, 0.46 and 0.55, respectively. The average digestible energy content
was 11.7 MJ/kg DM and average apparent ileal digestibility of essential amino
acids was 0.56.
It was concluded that, under tropical
conditions, the low levels of inclusion of leaves from mature cassava plants may
be useful in diets for growing pigs to improve the dietary protein supply.
Sun-drying and ensiling with cane molasses as additive successfully preserved
the nitrogenous constituents and could be a means of preserving fresh green
feed.
Keywords:
Pigs, rats, cassava leaves, amino acids, nitrogen utilization,
digestibility,
HCN.
Introduction
In
view of the predicted world shortage of cereal grains because of competing needs
for the expanding human population, the availability and supply of grains and
protein feedstuffs for livestock is likely to become more limited (Close 1993),
and more research into the use of non-conventional sources of energy and protein
for livestock production has been called for.
Cassava
(Manihot esculenta Crantz) is a widely grown crop in most countries in
the tropical regions of Africa, Latin America and Asia, and ranks as one of the
main crops in the tropical countries (Calpe1992). Cassava is extremely reliable
to grow, especially on sloping rain-fed soils of low fertility, survives drought
periods and grows well with limited supplies of water. In addition, it is
tolerant of acid soils and yields well on marginal soils without excessive use
of costly inputs. These qualities have endeared cassava to resource-poor
farmers.
Considerable
amounts of cassava leaves are readily available as a by-product at the time of
harvesting the roots. However, in the rainy season it is difficult to sun-dry,
and extending the drying period diminishes the nutritional quality of the
product. Ensiling could be a suitable alternative way of preserving the leaves.
Numerous reports have shown that cassava leaf has a high but variable protein
content (170 to 400 g/kg on a dry matter basis), with almost 0.85 of the crude
protein fraction as true protein (Ravindran 1993). While cassava leaf protein is
low in sulphur amino acids (Gomez and Valdivieso 1984), the content of most
other essential amino acids is higher than in soya bean meal (Eggum 1970). The high protein content and a
relatively good profile of essential amino acids are reasons for believing that
cassava leaves could be a potential protein source for monogastric animals.
Chemical composition of cassava leaves
The present studies have
demonstrated that there is considerable variation in the chemical composition
among leaves of different cassava varieties (Table 1), which is in agreement
with earlier reports (Ravindran 1990; Eggum 1970). The variation could be due to
differences in plant development as well as in the ratio of leaf/stem. The leaf
products were characterized by a higher crude protein content as compared with
the stems, while the NDF contents of the leaves were correspondingly lower.
Therefore, a further separation of the leaves from the structural plant elements
would probably have resulted in an upgrading of their nutritive value.
|
Table 1.
Nutrient content (g/kg DM) of leaves of some varieties of cassava grown in
South
Vietnam | |||||
|
Variety
/ source |
Crude
protein |
Ether
extract |
Crude
fibre |
NFE |
Ash |
|
KM94 |
347 |
133 |
120 |
338 |
62 |
|
India |
310 |
141 |
117 |
317 |
78 |
|
Gon |
285 |
135 |
146 |
355 |
79 |
|
Japan |
271 |
146 |
101 |
421 |
61 |
|
KM60 |
254 |
144 |
97 |
455 |
50 |
|
KM95 |
239 |
156 |
107 |
439 |
59 |
The cassava leaf products investigated in the present study were found to be high in crude protein (21 to 34 % of DM), and had an acceptable amino acid profile (Table 2), in comparison with, for example, alfalfa and soybean meal (Figure 1) (Phuc and Lindberg 2001). The contents of threonine in cassava leaves were quite acceptable compared with the ideal amino acid pattern for pigs, while the concentration of methionine and cystine, would be limiting (NRC 1998). The amino acid composition of cassava leaves is therefore of great importance when they are used as protein sources for monogastric animal species.
|
Table 2.
Chemical composition (% of DM), and
essential amino acid (EAA) and
non-essential amino acid (NEAA) content (g
per 16 g N) and energy content (MJ/kg DM)
in two batches of cassava leaves |
|||||
|
|
Batch
A |
|
Batch B
|
||
|
|
Sun
dried |
Dried |
Dried |
Sun
dried |
Ensiled |
Chemical composition
|
|
|
|
|
|
|
Organic matter |
92.8 |
92.4 |
92.8 |
91.4 |
92.8 |
|
Crude protein |
32.4 |
32.7 |
32.2 |
33.3 |
31.7 |
|
Ether extract |
6.4 |
7.5 |
8.2 |
7.2 |
8.1 |
|
NDF |
27.5 |
25.3 |
37.6 |
24.4 |
24.8 |
|
Gross
energy |
- |
- |
- |
21.4 |
21.2 |
Essential amino acids
|
|
|
|
|
|
|
Arginine |
6.3 |
6.4 |
5.8 |
6.5 |
5.6 |
|
Histidine |
2.2 |
2.0 |
2.0 |
1.8 |
1.7 |
|
Isoleucine |
4.1 |
4.4 |
4.5 |
4.2 |
4.2 |
|
Leucine |
8.7 |
8.9 |
9.1 |
8.3 |
8.3 |
|
Lysine |
5.1 |
5.1 |
4.2 |
5.5 |
5.4 |
|
Methionine |
1.6 |
1.4 |
1.5 |
1.6 |
1.4 |
|
Phenylal. |
6.3 |
6.2 |
6.2 |
6.2 |
5.6 |
|
Threonine |
4.4 |
4.2 |
4.4 |
4.1 |
3.9 |
|
Tyrosine |
4.3 |
4.6 |
4.6 |
4.4 |
4.4 |
|
Valine |
5.9 |
5.6 |
5.7 |
5.6 |
5.3 |
|
S EAA |
48.9 |
48.8 |
48.0 |
48.2 |
45.8 |
|
Non-essential
amino acids |
|
|
|
||
|
Alanine |
6.3 |
6.5 |
6.4 |
6.0 |
6.4 |
|
Aspartic acid |
9.7 |
9.4 |
9.6 |
10.3 |
9.3 |
|
Glutamic
“ |
11.0 |
10.4 |
10.7 |
9.3 |
9.6 |
|
Glycine |
4.5 |
4.4 |
4.8 |
4.3 |
4.1 |
|
Proline |
3.8 |
3.7 |
3.6 |
4.1 |
4.3 |
|
Serine |
3.4 |
4.1 |
4.2 |
3.3 |
3.8 |
|
S NEAA |
38.7 |
38.5 |
39.3 |
37.2 |
37.5 |
|
S AA |
87.6 |
87.3 |
87.3 |
85.5 |
83.3 |
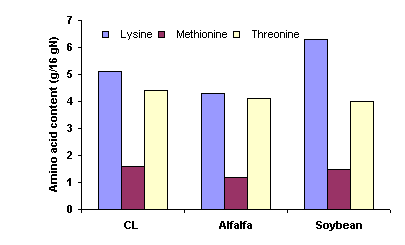
Figure
1. Amino acid contents of
cassava leaves (CL), alfalfa and soybean meal
Effect of inclusion of
cassava leaves on digestibility and nitrogen retention
Our studies have clearly demonstrated that increasing levels of inclusion of cassava leaves in diets for rats, as well as for growing pigs, lowered the ileal and total tract digestibility of organic matter and crude protein (Table 3). Figure 2a,b demonstrates the relationship between the dietary fibre (NDF) content and digestible organic matter for rats and pigs (Phuc and Lindberg 2000; Phuc et al 2001a).

Figure
2a: Effect
of level of dietary NDF on digestibility of
organic matter in rats
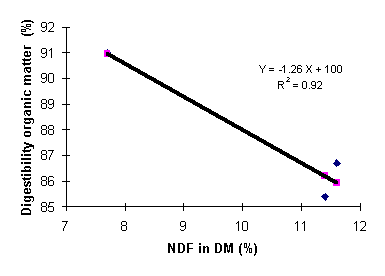
Figure
2b: Effect
of level of dietary NDF on digestibility of
organic matter in pigs
This can be explained by differences in chemical composition between the basal diet and the leaves, and the depressive effect of fibrous constituents on the apparent digestibility of the non-fibrous components of the diet (Stanogias and Pearce 1985a,1985b; Fernandez and Jřrgensen 1986). The negative effects of dietary fibre are partly a result of a reduced transit time of the ingesta in the small intestine caused by fibrous components, limiting the time for nutrient digestion and absorption (Den Hartog et al 1985; Sarwat et al 1986;). The extent of the reduction in digestibility has been shown to vary with the level of fibre and the feeding level (Oke 1990; Dierick et al 1989; Ravindran 1990). Furthermore, anti-nutritional factors in the cassava leaf products may have interfered with the process of digestion (Makkar 1993). Tannins are known to occur in cassava leaves (Ravindran 1993; Oke 1994), which may have adversely affected digestibility of crude protein.
Our study with pigs (Phuc
and Lindberg 2000) has shown that part of the dietary fibre had been digested at
the terminal ileum and that this slightly increased with the incorporation of
cassava leaves in the diets (Table 3). This suggests that the pigs were able to
digest a substantial part of plant fibre pre-caecally, which is in agreement
with earlier reports (Dierick 1989; Andersson and Lindberg 1997a,1997b).
Effects of inclusion of cassava leaves on
digestibility of AA at the terminal ileum
Most of the amino acids were
digested to a greater extent than the crude protein (Phuc and Lindberg 2001).
The reduction in apparent ileal digestibility of essential amino acids when
cassava leaves were included in the diets was due to an increase in the ileal
flow of amino acids with increasing fibre content in the diets, as reported by
Reverter et al (1999). Furthermore, an increase of endogenous amino acids
secretions could also be expected (Sauer and Ozimek 1986; Boisen and Moughan
1996; Jondreville et al 2000). The
magnitude of this effect will depend on the digestibility of the basal diet, the
level and type of the fibre and the contribution of the fibre source to the
dietary amino acid supply (Lenis et al 1996). In addition to fibre level and
quality, the presence of tannins and enzyme inhibitors (Kidder and Manner 1978)
might also influence the digestibility of protein and amino acids. The estimated
apparent ileal digestibilities of histidine, lysine and tyrosine were found to be highest,
while that of threonine was the lowest (Table 4).
Also the low arginine
digestibility of cassava leaves could be due to the binding of this amino acid
to lignin (Nongyao et al 1991). The average apparent ileal amino acid digestibility was 0.56, close to the
value reported for perennial ryegrass meal (0.61) (Reverter et al 1999).
|
Table
3. Estimated ileal and total tract
digestibility coefficients of nutrients and energy (%) in cassava leaves
in pigs | ||||||||
|
|
Ileal digestibility |
Faecal
digestibilty | ||||||
|
|
sun-dried |
ensiled |
sun-dried |
ensiled | ||||
|
|
Mean |
s.d |
Mean |
s.d |
Mean |
s.d |
Mean |
s.d |
|
Organic
matter |
0.41a |
0.03 |
0.42
a |
0.04 |
0.54
a |
0.03 |
0.59
a |
0.05 |
|
Crude protein
|
0.37 |
0.02 |
0.37 |
0.03 |
0.45 |
0.04 |
0.46 |
0.02 |
|
Crude
fat |
0.34
a |
0.02 |
0.41
b |
0.03 |
0.36
a |
0.02 |
0.43
b |
0.03 |
|
NFE
|
0.38 |
0.06 |
0.40 |
0.09 |
0.65 |
0.09 |
0.60 |
0.09 |
|
Crude
fiber |
0.34
a |
0.06 |
0.37
a |
0.04 |
0.50
a |
0.03 |
0.59
b |
0.03 |
|
NDF |
0.26
a |
0.04 |
0.23
a |
0.02 |
0.32
a |
0.02 |
0.31
a |
0.02 |
|
ADF |
0.16
a |
0.02 |
0.17
a |
0.02 |
0.20
a |
0.03 |
0.21
a |
0.04 |
|
Energy |
0.40
a |
0.06 |
0.41
a |
0.07 |
0.52
a |
0.04 |
0.57
b |
0.05 |
|
ab Means with different superscripts within rows are
significantly different
(P<0.05) | ||||||||
|
Table 4.
Estimated apparent ileal
digestibility of crude protein, and essential and non-essential amino
acids in sun-dried and ensiled cassava leaves | ||||
|
|
Sun-dried |
Ensiled | ||
|
|
Mean |
s.d |
Mean |
s.d |
|
Crude protein |
0.37 |
0.02 |
0.37 |
0.03 |
|
Essential amino acids |
|
|
| |
|
Arginine |
0.50a |
0.07 |
0.56 a |
0.05 |
|
Histidine |
0.61
|
0.05 |
0.68
|
0.03 |
|
Isoleucine |
0.48 a |
0.02 |
0.45 a |
0.04 |
|
Leucine |
0.50 a |
0.03 |
0.57
a |
0.02 |
|
Lysine |
0.64
a |
0.04 |
0.64 a |
0.04 |
|
Methionine |
0.56
a |
0.03 |
0.55 a |
0.04 |
|
Phenylalanine |
0.55 a |
0.05 |
0.52 a |
0.06 |
|
Threonine |
0.52 a |
0.07 |
0.54 a |
0.02 |
|
Tyrosine |
0.64 |
0.06 |
0.61 |
0.03 |
|
Valine |
0.60 a |
0.05 |
0.62 a |
0.03 |
|
Non-essential amino acids |
|
| ||
|
Alanine |
0.56 a |
0.02 |
0.57 a |
0.04 |
|
Aspartic
acid |
0.60 a |
0.04 |
0.62 a |
0.03 |
|
Glutamic
acid |
0.55 a |
0.08 |
0.53 a |
0.04 |
|
Glycine |
0.50 a |
0.06 |
0.54 a |
0.08 |
|
Proline |
0.62 a |
0.04 |
0.57 a |
0.07 |
|
Serine |
0.62 a |
0.03 |
0.61 a |
0.04 |
|
ab Means with different superscripts within rows
are different
(P<0.05) | ||||
Effect of inclusion of cassava leaves on
N retention and weight gains
The utilization of N
deteriorated with increasing inclusion of
cassava leaves. The most likely explanation for this is a low
digestibility of cassava leaves protein and a poorer amino acid profile. In
addition, the protein of cassava leaves is, compared with the basal diet,
deficient in the sulphur-containing amino acids (Eggum 1970; Yeoh and Chew
1976), and these may be used in the detoxification of HCN (Oke 1978, Ravindran
1993). This could further have imposed limitations on the efficiency of amino
acid utilization from HCN-containing cassava leaves in the present study.
Stepwise regression analyses indicated that methionine as well as lysine were
the major limiting amino acids for N retention as well as biological value (Phuc
et al 2001). The cassava leaf-based diets generally resulted in inferior weight
gains of the animals compared with the control diet, despite calculated
isoenergetic feed supply.
Effect of preservation techniques on
chemical composition of cassava leaves
Ensiled
cassava leaves tended to have higher digestibility coefficidents than sun-dried
leaves (Table 3), but differences were only significant for crude
fat. Sun-drying had only a small
effect on the crude protein content compared to the 60oC oven- drying
of CLM, which is in agreement with earlier studies (Ravindran et al.1987).
However, after drying at 105 oC Maillard reactions occurred and
lysine content was clearly lowered in CLM105 (4.2 g per 100 g crude protein
versus a value of about 5 g in the other cassava treatments, Table 2), most
likely due to the formation of lignin-like polymers, which also resulted in a
higher fibre content (Van Soest and Mason 1991). Therefore, rats fed diets with
inclusion of cassava leaves dried at 105 oC had clearly reduced N
utilisation as well as weight gain as a result of the lower digestible protein
and lysine content (Phuc et al 2001b).
Ensiling had only a limited effect on the
content of EAA as compared with drying (Table 2). The high content of lactic
acid (83.6 g/kg) and low content of ammonia (3.5 g/100 g N) as a percentage of
total N also confirmed the ideal ensiling conditions (Van Soest 1994) resulting
from cane molasses addition (Phuc et al 2001b). However, the unpleasant taste
and/or smell related to the low pH and the occurrence of organic acids in the
ensiled products were possibly the cause of the reduced feed intake in rats. The
negative influences on digestibility of organic matter, crude protein and N
utilisation, which also reduced the weight gain of the rats, were observed when
the silage cassava leaves were
included in the diet.
Hydrogen cyanide (HCN)
The HCN content varied among
the varieties, from 285 to 762 mg/kg DM. It seems that the slow drying rate
achieved by sun-drying resulted in an increase of cyanogenic activity, thereby
eliminating cyanides as compared with the fast artificial drying at 60
oC, which is in line with earlier work (O’Brien and Jones 1994) (Phuc
et al 2001). However, at 105 oC the drying temperature had a marked
effect on the cyanide content of cassava leaves and their consequent toxicity (Ravindran
1993; Nambisan 1994; Oke 1994). The higher total HCN content in cassava leaf
silage as compared to cassava leaf meal (Table 5) is confirmed by other studies
(Gomez and Valdiviesi 1988; Oke 1994; Westby 1994).
|
Table
5. Contents
of total HCN, intermediary products
(cyanohydrine), free HCN and glucoside (linamarin)
(mg/kg DM) of cassava leaves |
|||||
|
|
Cassava
leaves
A |
Cassava
leaves B
|
|||
|
|
Sun
dried |
Dried
60 oC |
Dried
105 oC |
Sun
dried |
Ensiled
dried |
|
Total HCN |
59 |
86 |
28 |
255 |
250 |
|
Cyanohydrine |
13 |
47 |
1 |
152 |
215 |
|
Free HCN |
13 |
33 |
9 |
62 |
5 |
|
Linamarin |
33 |
6 |
18 |
42 |
30 |
|
HCN
content of ensiled cassava leaves (not
dried): total HCN: 762; Cyanohydrine: 673;
Free HCN: 17 and Linamarin: 73. |
|||||
It can be concluded that
drying and ensiling are effective ways of reducing the toxicity of cassava
products, which is in agreement with the conclusion of Twe (1991). There were no
indications of cyanide toxicity on any of the diets in the present study in the
rat as well as the pig experiments, for both sun-dried and ensiled cassava
leaves (Phuc et al 2000; Phuc et al 2000a,b; Phuc and Lindberg 2001). This
finding contrasts with the recommended safe level of HCN in the diet, which is
below 50 mg HCN/kg according to Bolhuis (1954), a level which was not achieved
in any of the diets tested.
Estimation of the nutritive value of cassava leaves in rats and pigs
The chemical composition of
the cassava leaves tested with rats and pigs and their nutritive value estimated
by the difference and regression methods are shown in Table 6. The present data
do not allow testing of the hypothesis that estimates of the nutritive values
assessesed with rats are applicable to pigs, as the chemical composition of
cassava leaves used for the two animal species were different, and also because
the number of products tested was too limited.
|
Table 6.
Energy (GE, MJ per kg DM), crude protein (CP) and NDF (% of DM),
digestibility coefficients (%) and estimated digestible energy content
(DE, MJ/kg DM) of cassava leaves investigated in pigs and rats | |||||||
|
|
CLM |
ECL | |||||
|
|
Rats |
Pigs |
Rats |
Pigs | |||
|
|
I |
I
|
II |
II |
I |
III |
IV |
|
Chemical
composition |
|
|
|
| |||
|
GE |
21.3 |
19.2 |
21.4 |
- |
18.4 |
21.2 |
- |
|
OM |
92.8 |
91.4 |
91.3 |
89.1 |
92.8 |
89.5 |
89.7 |
|
CP |
32.4 |
33.3 |
26.4 |
26.0 |
31.7 |
24.5 |
27.6 |
|
NDF |
24.5 |
27.4 |
32.1 |
33.5 |
24.8 |
32.6 |
33.5 |
|
Digestibility |
|
| |||||
|
OM |
59 |
59 |
54 |
51 |
60 |
59 |
52 |
|
CP |
61 |
59 |
45 |
44 |
62 |
46 |
59 |
|
DE |
11.5 |
11.3 |
11.0 |
- |
11.0 |
12.0 |
- |
|
I Phuc et al 2001a; II
Phuc et al 2001b; III. Phuc and Lindberg 2000; IV Phuc et al 2000 | |||||||
Practical
considerations
The present studies have
demonstrated that growing pigs have the capacity to utilize the nutrients and
energy of cassava leaves. However, the potential benefits of an early harvest of
the leaves (eg: when it is managed as a forage; Preston 2001) should be
stressed. A separation of leaves and stems could further improve the nutritional
properties of the cassava biomass, and this would be more critical for pigs. As
the capacity of the pig to digest fibrous components improves as the pig matures
(Kenneth et al 1990) this strategy would be most beneficial for the nutrient
supply of piglets, young growing pigs and lactating sows, which have high demands for nutrients.
Cassava leaves contain more
protein and amino acids per energy unit than cereal grains, thereby also
reducing the need for protein supplements. However, the content of crude protein
in cassava leaves as well as its digestibility, is much lower than that of
soybean meal or other traditional protein supplements. Therefore, it is
necessary to determine the extent to which they can contribute to the supply of
nutrients for different categories of animals, as well as their nutritional
limitations, especially antinutritional factors. If these are known, feeding
strategies based on cassava leaves could be developed that would provide an
adequate supply of both energy and protein to meet the animals’ nutritional
requirements.
References
Andersson C
and Lindberg J E 1997a Forages in diets for growing pigs. 1. Nutrient apparent
digestibilities and partition of nutrient digestion in barley-based diets
including lucerne and white-clover meal.
Anim. Sci. 65:
483-491.
Andersson C
and Lindberg J E 1997b Forages in diets for growing pigs. 2. Nutrient apparent
digestibilities and partition of nutrient digestion in barley-based diets
including red-clover and perennial ryegrass meal. Anim. Sci. 65:
493-500.
Boisen S
and Moughan P J 1996 Difference expressions of dietary protein and amino acid
digestibility in pig feeds and their application in protein evaluation: A
theoretical approach. Acta Agriculturś
Scandinavica, Section A, Anim. Sci. 46: 165-172.
Bolhuis G G 1954 The toxicity of cassava root. J. Agric. Sci. 2:
176185.
Calpe C 1992 Roots, tubers and plantains: Recent trends in production, trade and use. In: Machin D, Nyvild S (Eds) Roots , Tubers, Plantains and Bananas in Animal Feeding. FAO Anim. Prod. and Health Paper 95: 11-25
Close W H
1993 Fibrous diets for pigs. Anim. Prod. in Developing Countries.
16,107-117.
Den Hartog
L A, Verstegen M W A,
Huisman J and van Kempen G J M 1985 Effect of dilution of a pig diet on the digestibility of
the nutrients. Proc. of 36th
annual meeting of the Eur. Assoc. for Anim. prod. (EAAP). Halkidiki, Greece.
Dierick N
A, Vervaeke I J, Demeyer D I and Decuypere J A 1989 An approach to the energetic importance of fibre
digestion in pigs. I. Importance of fermentation in the overall energy supply.
Anim. Feed Sci. Technol. 23:
141-167.
Eggum B O 1970 The protein quality of cassava leaves. Br. J. Nutr. 24:
761-768
Fernández J A and Jřrgersen N
1986 Digestibility and absorption of nutrients
as affected by fiber content in the diet of the pig. Quantitative aspects. Livest. Prod. Sci. 15:
53-71.
Gomez G
and Valdivieso M 1984 Cassava for animal feeding: Effect of variety and plant
age on production of leaves and roots. Anim. Feed Sci. Technol.
11,49-55.
Jondreville C, Van Der Broecke J, Grosjean F, Van Cauwenberghe S
and Gatel F 2000 Ileal true digestibility of amino acids in wheat milling
by-products for pigs. Ann. Zootechnie. 49:
55-65.
Kenneth B
K, Gilbert R H and Murray Danielan D
1990 Forages for swine. In: Pork Industry Handbook. Purdue
University Cooperative Extention Service. West Lafayette, Indiana. USA. PIH-126.
7 pp
Kidder D E and Manner M J 1978 Digestion in the pig. Scientechnica Bristol. 201
pp.
Kumar R
and D‘Mello J P
F 1995 Anti nutritional
factors in forage legumes. In: Tropical
Legumes in Animal Nutrition (Eds): D‘Mello, P F
and Devendra C CAB. International, Wallingford, UK. pp.
95-133.
Lenis N P, Bikker P, van der Meulen J, van Diepen J Th. M, Bakker J G M and Jongbloed A W 1996 Effect of dietary Neutral Detergent Fiber on ileal
digestibility and portal flux of nitrogen and amino acids and on nitrogen
utilization in growing pigs. J. Anim.
Sci. 74:2687 – 2699.
Mahendranathan T
1971 The effect of feeding tapioca (Manihot
utilissima Pohl) leaves to pigs. Malays. Agric. J. 48:
60-68.
Makkar H P S 1993 Anti-nutritional factors in foods for livestock. Animal
Production in Developing Countries. Br.
Soc. Anim. Prod. 16: 69-85.
Nambisan B
1994 Evaluation of the effect
of various processing techniques on cyanogen content reduction in cassava. In:
Acta Horticulturea Cassava Safety
375. Proc. Int. Workshop in Nigeria.
pp.193-201.
Nongyao A, Han I K and Choi Y J 1991 Amino acid digestibility to pigs in various fiber
sources 2. True digestibility of amino acids in ileal digesta and feces. Asi. Austr. J. Anim. Sci. 4 (3):
211-218.
NRC 1998 Nutrient requirements of
swine. Tenth revised edition. Washington D.C
O’Brien G M and Jones D
M 1994 Processing approaches to optimizing raw materials and
end product quality in the production of cassava flours. In: Acta Horticulturea Cassava Safety 375.
Proc. Int. Workshop in Nigeria. pp.183-191
Oke O L 1990 Cassava meal. In:Thacker P A ; Kirlwood R N (Eds) Non-traditional feed source for
use in swine production. Butterworths Boston, 103-112
Oke O L 1994 Eliminating cyanogens from cassava through processing:
Technology and tradition. In: Acta
Horti.Cassava Safety 375. Proc. Int. Workshop in Nigeria. pp.163-174
Phuc B
H N and Lindberg J
E 2001 Ileal digestibility of
amino acids in growing pigs fed a cassava root meal diet with inclusion of
cassava leaves, leucaena leaves and groundnut foliage. Anim. Sci. 72:
511-517.
Phuc B
H N, Lindberg J
E, Ogle B and Thomke S
2001a Nutritive evaluation with rats of tropical biomass
products for monogastrics: 1. Comparison of eight green products evaluated at
two levels of inclusion in relation to a casein diet. Asian-Aust. J. Anim. Sci. (In
press).
Phuc B
H N, Lindberg J
E, Ogle B and Thomke S
2001b Nutritive evaluation with rats of tropical biomass
products for monogastrics: 2. Effects of drying temperature, ensiling and level
of inclusion of cassava leaves and sweet potato vines. Asian-Aust. J. Anim. Sci. (In
press).
Phuc B H N and Lindberg J E 2000 Ileal and total tract digestibility in growing pigs fed
cassava root meal diets with inclusion
of cassava leaves, leucaena
leaves and groundnut foliage. Anim.
Sci. 71:301-308.
Phuc B H N,
Ogle B and Lindberg J E 2000 Effect of replacing soybean meal with cassava leaf
protein in cassava root meal based diets for growing pigs on digestibility and N
retention. Anim. Feed Sci. Technol.
83: 223-235.
Ravindran V
1990 Cassava leaf meal. In: Non- Traditional Feed Sources for Use in
Swine Production. (Eds), Thacker
P A and Kirkwood R N Butterworths, Boston. pp 91-101.
Ravindran V
1993 Cassava leaves as animal feed: Potential
and limitations. J. Sci. Food Agric. 61:
141-150.
Reverter M,
Lundh T and Lindberg J E 1999 Ileal amino acid digestibilities in pigs of barley-based
diets with inclusion of lucerne, white clover, red clover or perennial ryegrass.
Br. J. Nutr. 82:
139-147.
Sarwat S V, Kakala S N , and Kategile J A 1998 Performance of growing-finishing pigs
when fed diets containing fresh cassava leaves and roots. East Afr. Agric. Forest. J 53(3): 111-115.
Sauer W C and Ozimek L 1986 Digestibility of amino acids in swine: results and their
practical applications. A review. Livest.
Prod. Sci. 15: 367 – 388.
Stanogias G
and Pearce G R 1985a The digestion of fibre by pigs. 1. The effects of amount
and type of fibre on apparent digestibility, nitrogen balance and rate of
passage. Br. J Nutr. 53:
513-530.
Stanogias G
and Pearce G R 1985b The digestion of fibre by pigs. 3. Effects of the amount
and type of fibre on physical characteristics of segments of the
gastrointestinal tract. Br. J. Nutr.
53: 537-548.
Twe O O 1991 Detoxification of cassava products and
effects of residual toxins on consuming animals. In: Roots, tubers, plantains and bananas in
animal feeding. (Eds) Machin ,D, Nyvild, S FAO Anim. Prod. Health Paper 95:
81-95.
Van
Soest P J 1994 Nutritional ecology of the ruminant.
2nd edition. Cornell University Press, Ithaca,
USA.
Van Soest P J and Mason V C 1991 The influence of the Maillard reaction upon the
nutritive value of fibrous feeds. Anim.
Feed Sci. Technol. 32: 45-53.
Westby A
1994 Cyanogen
reduction during lactic fermentation of cassava. In: Acta Horticulturea Cassava Safety 375.
Proc. Int. Workshop in Nigeria. pp.
209-215.
Yeoh H H and Chew M
Y 1976 Protein content and amino acid
composition of cassava leaf. Phytochem.
15: 1597-1599.