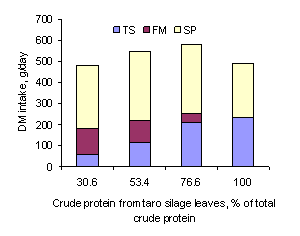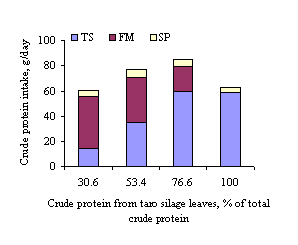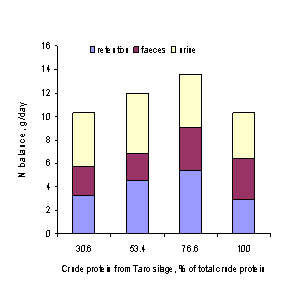The four treatments applied to 4 growing pigs
(15 kg live weight) in a 4*4 Latin square arrangement were four levels of
ensiled leaves of taro (Colocasia esculenta) equivalent to 25, 50, 75 and
100% substitution of the protein from fish meal in a basal diet of sugar palm
syrup. The pigs were housed in metabolism cages for consecutive periods of 12
days, with collection of faeces and urine during the last 5 days of each period.
There were positive curvilinear trends in DM and
crude protein intake and in N retention in response to replacement of fish meal
by ensiled taro leaves with the optimum proportion being at about 70-75%
replacement. Coefficients of DM apparent digestibility were high on all diets
but those for crude protein showed a negative curvilinear response with
declining values beyond 25% substitution of the fish meal protein by that from
taro leaf silage. The limiting factor to utilization of the taro leaf silage
appears to be the lower apparent digestibility of the protein in the taro
leaves.
Taro (Colocasia esculenta), also known as Old Cocoyam,is a
wetland crop in many tropical and subtropical areas of the world, cultivated
mainly for root (corm) production. Before the corm can be eaten, the traditional
practice is to cook it so as to break down the needle-like calcium oxalate
crystals which are present in all parts of the plant. According to Miller
(1929), these crystals are extremely irritating to the throat and mouth lining,
causing a burning and stinging sensation. Many other plants contain substantial
quantities of oxalate (Standal 1983). Proposed functions of the oxalate content
in plants are said to include: protectionfrom insects and foraging
animals through toxicity and/or unpalatability,osmo-regulation, and
regulation of calcium levels in plant organs andtissues (Libert and
Franceschi 1987).
In a survey in Takeo and Pursat provinces, 51% of the farmers that were
interviewed reported feeding the petiole of Taro (Paper I) to pigs. As for the
corm, the traditional practice is to boil the petioles before feeding. Very few
farmers reported feeding the leaves. In the mountain areas of North Vietnam,
there is a tradition of using both the corm and the leaves of Giant taro (Alocasia
macrorrhiza) as feed for pigs (Tiep et al 2006). As in the case of taro, it
was reported (Tiep et al 2006) that the traditional way is to boil the corm and
the leaves prior to feeding, as this reduced the concentration of calcium
oxalate present. These authors showed that ensiling the leaves with rice bran
and molasses reduced the calcium oxalate content by 78%, and that the silage
could be included in the diet of growing pigs at the 10% level without affecting
growth rate. In Colombia, Rodriguez et al (2006) showed that the leaves of the
New Cocoyam (Xanthosoma sagittifolium) could be fed fresh to young
pigs at levels replacing 50% of the soybean meal (approximately 45% of the diet
DM) and that growth rates (500 g/day) were similar to those on the control diet
without the leaves. The leaves were chopped before offering them to the pigs
which may have reduced the oxalate content as it was observed that the
irritating effect from handling the leaves was reduced after chopping them
(Preston TR personal communication).
There appears to be little recent research on the feeding value of taro leaves
for pigs. For this reason the present study was carried out to provide
preliminary information on digestibility and nitrogen balance in pigs fed
ensiled taro replacing fish meal, using a fibre-free basal diet (sugar palm
syrup), as according to Preston (2006) this is likely to facilitate the use of
fibrous protein sources, such as leaves from vegetative sources.
Materials and methods
Location and Climate
The experiment was conducted in the ecological farm of the Center for Livestock
and Agriculture Development (CelAgrid), located in Rolous village, Rolous
commune, Kandal Stoeung district, Kandal province, about 26 km from Phnom Penh
City, Cambodia. During the trial (from 1 September to 21 November, 2006), the
average ambient temperature was 25.9± 0.96 C in the morning at
06:00h, 31.7 ± 1.59 C in the middle of day, and 28.3 ± 1.90 C
in the afternoon at 18.00h.
Experimental design
The experiment was carried out as a Latin Square 4x4 arrangement with 4 pigs and
4 periods (Table 1), each of 12 days. The first 7 days were for adaptation to
the diet, followed by 5 days for collection of faeces and urine.
Table 1.
Experimental layout of the digestibility trial
Period
Pig 1
Pig 2
Pig 3
Pig 4
1
TS25
TS50
TS75
TS100
2
TS100
TS75
TS50
TS25
3
TS75
TS100
TS25
TS50
4
TS50
TS25
TS100
TS75
Animals and housing
Four crossbred (Large White x Local) castrated male pigs,
initially averaging 15.0 ± 1.0 kg live weight were used for the experiment. They
were housed in metabolism cages (1.6 x 1.4 m floor area), elevated 0.6 m and
allowing the separate collection of faeces and urine. The cages were made of
wood and were situated in a building with a thatch roof of Imperata grass, and
open sides.
Diets and feeding
management
Wild taro (Trav Prey) leaves were harvested from a lake receiving waste water
from Phnom Penh City. The leaves were chopped into small pieces and ensiled with
5% of sugar palm syrup (79% of DM) and 2% of water (fresh basis) to facilitate
the mixing of the viscous syrup. Leaves and syrup (plus the water) were
carefully mixed and stored in plastic bags, inside rigid plastic containers. The
bags were tightly closed to prevent air contact and stored for 30 days before
feeding. Every container thus prepared contained approximately 150 kg of fresh
material. The ensiled Taro leaves had a pH of
3.85 after one month and DM content of 18.3%.
Four diets were formulated with protein from ensiled
taro leaves replacing fish meal protein at levels of 25, 50, 75 and 100% (Table
2). The pigs were fed ad libitum, with fresh feed supplied 3 times
per day during the adaptation period, and given 80% of the mean ad libitum
intake in the collection period to avoid residues. All ingredients were mixed
together before feeding. Fresh water was available at all times from nipple
drinkers. The duration of the experiment was 48 days.
Table 2.
Chemical composition of ensiled taro leaves and ingredient and
chemical composition of the experimental diets (% in dry matter)
Ensiled taro leaves
Fish meal
Sugar palm syrup
Replacement of
fish meal protein, %
25
50
75
100
Ingredients,%
Ensiled taro leaves
-
-
-
11.5
23
34.5
46
Fish meal
-
-
-
25.0
16.8
8.4
0
Sugar palm syrup
-
-
-
62.0
58.7
55.6
52.5
Salt
-
-
-
0.5
0.5
0.5
0.5
Premix (mineral-vitamin)
-
-
-
1.0
1.0
1.0
1.0
Analysis
Data calculated from analysis of
the ingredients
Dry matter %
18.3
92.1
79
75.4
67.4
59.3
51.2
As % of DM
Ash
8.3
56.8
0.02
15.2
11.5
7.63
3.81
Crude protein
25.9
35.6
0.24
12
12
12
12
Calcium oxalate
0.11
-
-
0.013
0.025
0.038
0.051
Measurements
Feeds offered and residues, and output of faeces and urine were recorded daily
during the last 5 days of each period. The animals were
weighed in the morning before feeding at the beginning and the end of
each period. Samples of each component (offered) were retained for analysis of
DM, Ash, OM and N. A sub-sample (10%) of faeces was put each day in plastic bags
in the freezer (-20 °C). Urine was collected in a bucket containing 10 ml of 10%
sulphuric acid (H2SO4) to keep the pH below 4 so as to
prevent escape of ammonia. A sub-sample (10%) was collected each day and kept in
the freezer.
Chemical analysis
Feed and faecal samples were dried by microwave radiation to measure the DM
content (Undersander et al., 1993). Total N of samples of feed and faeces and
urine was determined by the Kjeldahl procedure as outlined in AOAC (1990). The
ash content of feed and faeces was determined following the AOAC (1990)
recommendations; organic matter was assumed to be the result of subtracting the
percentage of ash from 100. The calcium oxalate was determined according to AOAC
(1990) procedures.
Statistical analysis
The data were analyzed using the general linear model (GLM) procedure in the
ANOVA software of MINITAB (release 13.31). Sources of variation were treatments,
periods, animals (pigs) and error.
Results and discussion
The process of ensiling the taro leaves was more effective than sun-drying in
reducing the calcium oxalate content (Table 3). The concentration in the ensiled
leaves was lower than reported by Tiep et al (2006) (0.7% in DM) for ensiled
leaves of Giant taro (Alocacia macrorrhiza). The fish meal used was of
extremely poor quality, with a crude protein content of only 35.6% of DM, and an
ash content of 56.8%, probably because the meal was contaminated, possibly by,
for example, addition of sand.
Table 3.
Effect of sun-drying and ensiling on calcium oxalate in the taro
leaves, % in DM
Fresh
Sun-dried
Ensiled
3.08
1.10
0.11
There were slight differences between the planned and observed levels of
substitution of fish meal protein by the
protein from taro leaves, with higher than intended levels for the intermediate
treatments TS25, TS50, and TS75 (Table 4). This was because of a small
degree of selection by the pigs.
Table 4.
Proportions (%) of the protein supplement provided by taro leaves
and fish meal; planned and
recorded in the experiment
TS25
TS50
TS75
TS100
Planned
25
50
75
100
Recorded
30.6
53.4
76.6
100
Daily feed intake
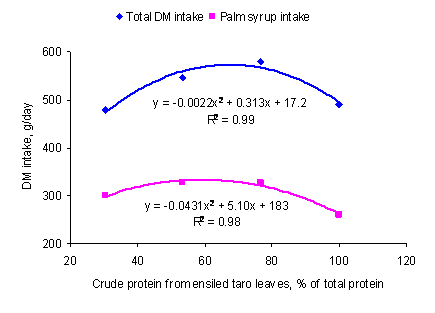
There were significant differences between treatments for intake of palm syrup
and total DM (Table 5). The relationship between DM intake and dietary protein
from ensiled taro leaves was curvilinear (Figure 1) with maximum intake at about
the 75% replacement of fish meal, followed by a decline to the 100% level. The
trend for intake of palm syrup was similar. The reason for the decrease in
intake of the TS100 diet may have been the bulkiness of the diet because of
the high content of the low DM silage. The DM intake expressed as a function of
live weight on the TS100 diet (30.1 g/kg) was lower than reported by Rodriguez
et al (2006) (48.5 g DM/kg live weight) for a similar diet fed to pigs of
similar live weight, but with sugar cane juice instead of palm syrup and fresh
leaves of Xanthosoma sagittifolium instead of ensiled leaves of
Colocasia esculenta.
Table 5.
Mean values for feed and nutrient intake in pigs fed sugar palm
syrup with increasing proportions (25 to 100%) of protein from
ensiled taro leaves replacing protein from
fish meal
TS25
TS50
TS75
TS100
SEM
P
DM intake,
g/day
Taro silage
60.7a
118b
208c
232c
Fish meal
118d
100c
46.4b
0a
Sugar palm
syrup
301b
328b
327b
261a
9.10
0.001
Total DM
480a
546b
581b
491a
15.50
0.001
DM, g /kg LW
24.9a
34.1b
33.2b
30.1b
2.35
0.032
Figure 1:
Relative intakes of DM from dietary ingredients (TS taro leaf silage, FM
fish meal, SP sugar palm syrup)
Figure 2:
Relative intakes of crude protein from dietary ingredients
Figure3.
Relationship between intake of palm syrup and total DM and
proportion of dietary protein from ensiled taro leaves
Apparent digestibility
There were no significant differences among treatments in apparent digestibility
of DM and organic matter (Table 6) which were high in all cases. The value for
DM of 87.7 on the TS100 diet was slightly higher than was reported by Rodriguez
et al (2006) for a diet of sugar cane juice and fresh leaves of Xanthosomasagittifolium (83.4%). If it is assumed that the sugar palm syrup is 100%
digested, and as it contributed 53% of the diet DM in treatment TS100, it can be
predicted that the apparent digestibility of the taro leaf silage would be 73.8%
for the DM and 86% for the organic matter.
Table 6.
Mean values for diet digestibility in pigs fed sugar palm syrup with
increasing proportions (25 to 100%) of protein from taro leaf silage
replacing protein from fish meal
TS25
TS50
TS75
TS100
SE
P
Apparent
digestibility, %
Dry matter
84.7
85.6
84.5
87.7
1.4
0.370
Crude protein
75.3ab
78.1b
69.4ab
66.1a
2.8
0.019
OM
89.6
91.5
88.6
93.4
2.13
0.418
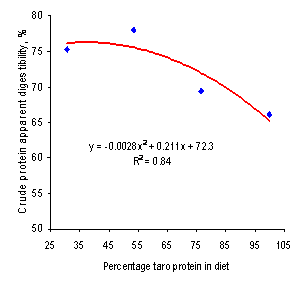
Crude protein apparent digestibility differed among treatments (P<0.05) with a
negative curvilinear trend (Figure 4), as the proportion of protein from taro
silage was increased, with the lowest value of 66.1% on the TS100 diet. A
similar negative trend was reported by Rodriguez et al (2006) with a value of
61.4% for the diet with only cane juice and Xanthosoma leaves. These
negative effects can be attributed to the binding of some of the proteins to the
fibre which reduces its digestibility (Jørgensen et al 1996; Kass et al 1980).
Shayo and Uden (1999) reported that 64% of the protein in Xanthosoma
leaves was in the NDF fraction; however, a much smaller fraction (21%) was
reported by Leterme et al (2005) for leaf protein in Xanthosoma.
Figure 4.
Relationship between apparent digestibility of crude protein and
proportion of diet protein from ensiled taro leaves
Figure 5.
Mean values for N balance
in pigs fed sugar palm syrup with increasing proportions of
protein from taro leaf silage replacing protein from fish meal
Nitrogen balance
There were significant differences in all the components of N balance (Table 7;
Figure 5). Faecal N tended to increase (P<0.05) and urine N to decrease as the
proportion of taro leaf silage increased. Similar trends were reported by
Rodriguez et al (2006) for diets of sugar cane juice and Xanthosoma
leaves. There were significant effects of N intake on N retention (P<0.001), but
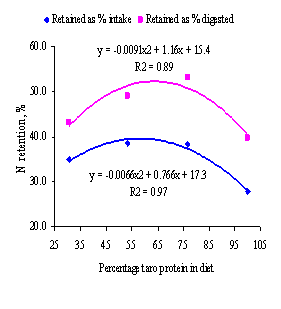
not on N retention as % of intake and % of digested N (P>0.05). The responses to
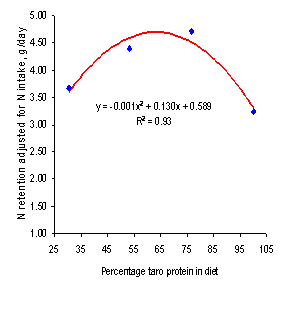
increasing intake of dietary taro leaf protein were curvilinear for N retention
(adjusted for N intake; Figure 6) and for N retained as % of N intake and as %
of digested N (Figure 7). This response followed closely the trend for DM
intake (Figure 1) implying that the best results would be with 75% replacement
of the fish meal by the taro leaf silage. The similar values for N retention on
TS0 (all fish meal) and TS100 (all taro silage) suggests that the amino balance
was not a limiting factor to pig performance. This is supported by the
conclusion of Leterme et al (2005) that tropical tree leaves offer a variable
amount (170-240 g kg−1) of proteins that are well balanced in
essential amino acids but not well digested by pigs.
The N retention values in general were lower (range 2.9 to 5.4 g N/day) than
those recorded by Rodriguez et al (2006) (range 6.1 to 9.7 g N/day). The main
reason for this would seem to be the lower DM intake (25 to 34 g DM/kg LW)
compared with the experiment of Rodriguez et al (2006) (49 to 56 g DM/kg LW).
The diets were similar in overall composition (mainly sugars as the energy
component and leaves of similar plants species (Old Cocoyam versus New
Cocoyam)). The difference in response in feed intake could be due to the
previous feeding history of the two sources of pigs. These were of similar live
weight (15 kg in this study versus 13.4 kg in the experiment of Rodriguez et al
(2006)). However, the pigs in the latter study were from sows that had been fed
leaves of New Cocoyam during pregnancy and lactation, when the piglets also had
access to the leaves. Tien and Preston (2003) concluded that reductions in
growth rate at weaning time when piglets are first offered solid foods, or
subsequently due to frequent changes in diet, can be reduced if novel foods are
eaten pre-natally by mother or post-natally by mother and piglets together.
Table 7.Mean values for N balance in
pigs fed sugar palm syrup with increasing proportions of protein
from taro leaf silage replacing protein from
fish meal
TS25
TS50
TS75
TS100
SEM
P-value
Components of N balance, g N/day
Intake
10.2a
12.0b
13.5c
10.4a
0.36
0.001
Faeces
2.47a
2.21b
3.68a
3.57a
0.33
0.019
Urine
4.53ab
5.18a
4.47ab
3.82b
0.33
0.053
N retained
g/day
3.26b
4.58a
5.42a
2.89b
0.33
0.001
g/day#
3.68
4.40
4.72
3.24
0.36
0.099
%
of N intake
34.9
38.4
38.2
27.7
3.00
0.13
%
N digested
43.2
49.1
53.1
39.6
3.80
0.42
# Adjusted
for differences in N intake by using
covariate analysis
Figure 6.
Relationship between N retention (adjusted for N intake) and
proportion of diet protein from ensiled taro leaves
Figure 7.
Relationship between N retention as % of intake and % of digested N,
and proportion of diet protein from ensiled taro leaves
Conclusions
There were positive curvilinear trends in DM and crude protein intake and in
N retention in response to replacement of fish meal by ensiled taro leaves
with the optimum proportion being at about 70-75% replacement.
Coefficients of DM apparent digestibility were high on all diets but those
for crude protein showed a negative curvilinear response with declining
values beyond 25% substitution of the fish meal protein by that from taro
leaf silage.
The limiting factor to utilization of the taro leaf silage appears to be the
lower apparent digestibility of the protein in the taro leaves
and maybe amino acid imbalance of the protein, or
a greater proportion of crude protein fermented in the caecum.
Acknowledgements
The present experiment is
part of a study on ensiled taro (Colocasia esculenta) leaves in diet of
local crossbred pigs in Cambodia, supported by the MEKARN project financed by
SIDA-SAREC. The authors express their gratitude to all the personal of the
CelAgrid Ecological Farm, for help with the experiment. This paper
forms part of the MSc thesis (MEKARN-SLU, Uppsala, May 2007) of the senior
author.
References
AOAC 1990. Official Methods of Analysis. Association of
Official Analytical Chemists. 15th edition (K Helrick, editor). Arlington pp
1230
Jørgensen H, Zhao X, Eggum B O and Zhao X Q
1996: The influence of dietary fiber
and environmental temperature on the development of the gastrointestinal tract,
digestibility, degree of fermentation in the hind-gut and energy metabolism in
pigs. British Journal of Nutrition 75, 365-378. http://cat.inist.fr/?aModele=afficheN&cpsidt=3032927
Kass M L, Van Soest P J, Pond W G, Lewis B
and McDowell R E 1980: Utilization of
dietary fiber from alfalfa by growing swine. I. Apparent digestibility of diet
components in specific segments of the gastrointestinal tract.Journal
of Animal Science50, 175-181.
Libert B and Franceschi VR 1987.
Oxalate in crop plants. J Agric Food Chem 35: 926-938
Miller CD 1929.
Food Values of Breadfruit, Taro Leaves, Coconut and Sugar Cane.
Tien D V and Preston T R 2003.
Pre and post-natal exposure to duckweed affects its post-weaning familiarity and
intake in Large White and Mong Cai pigs. (Msc
Thesis in Sustainable Livestock Production, Swedish University of agricultural
Sciences, Uppsala). http://www.mekarn.org/sarec03/Tien2.htm
Tiep P S, Nguyen Van Luc, Trinh
Quang Tuyen, Nguyen Manh Hung and Tran Van Tu 2006. Study on the use of Alocasiamacrorrhiza(roots
and leaves) in diets for crossbred growing pigs under mountainous village
conditions in northern Vietnam. Workshop-seminar "Forages for Pigs and Rabbits"
MEKARN-CelAgrid, Phnom Penh, Cambodia, 22-24 August, 2006. Article # 11.
RetrievedFebruary 8, 107, from http://www.mekarn.org/proprf/tiep.htm
Preston TR 2006. Forages as protein sources for pigs in the tropics.
Workshop-seminar on Forages for Pigs and Rabbits MEKARN-CelAgrid, Phnom Penh,
Cambodia, 22-24 August, 2006.
Rodríguez L, Peniche I and
Preston T R 2006. Digestibility and
nitrogen balance in growing pigs fed a diet of sugar cane juice and fresh leaves
of New Cocoyam (Xanthosomasagittifolium) as partial or complete
replacement for soybean protein. Workshop-seminar "Forages for Pigs and Rabbits"
MEKARN-CelAgrid, Phnom Penh, Cambodia, 22-24 August, 2006. Article # 15.
RetrievedFebruary 8, 107, from http://www.mekarn.org/proprf/rodr2.htm
Shayo C and Uden P 1999.
Nutritional uniformity of crude protein fractions in some tropical browse plants
estimated by two in vitro methods. Anim Feed Sci Technol
78:141151.
Standal B R 1983.
Nutritive value. In JK Wang, ed, Taro: A Review of Colocasia esculenta
and Its Potentials. University of Hawaii Press, Honolulu, pp 141-147
Undersander D, Mertens D R and Thiex N
1993. Forage analysis
procedures. National Forage Testing Association. Omaha, pp 154.